L05 Photosynthesis
一、The Basic Processes of Photosynthesis
Photosynthesis:
Provides carbohydrates for energy production
Fixes CO2 (carbon re-enters the biosphere)
The major source of atmospheric O2
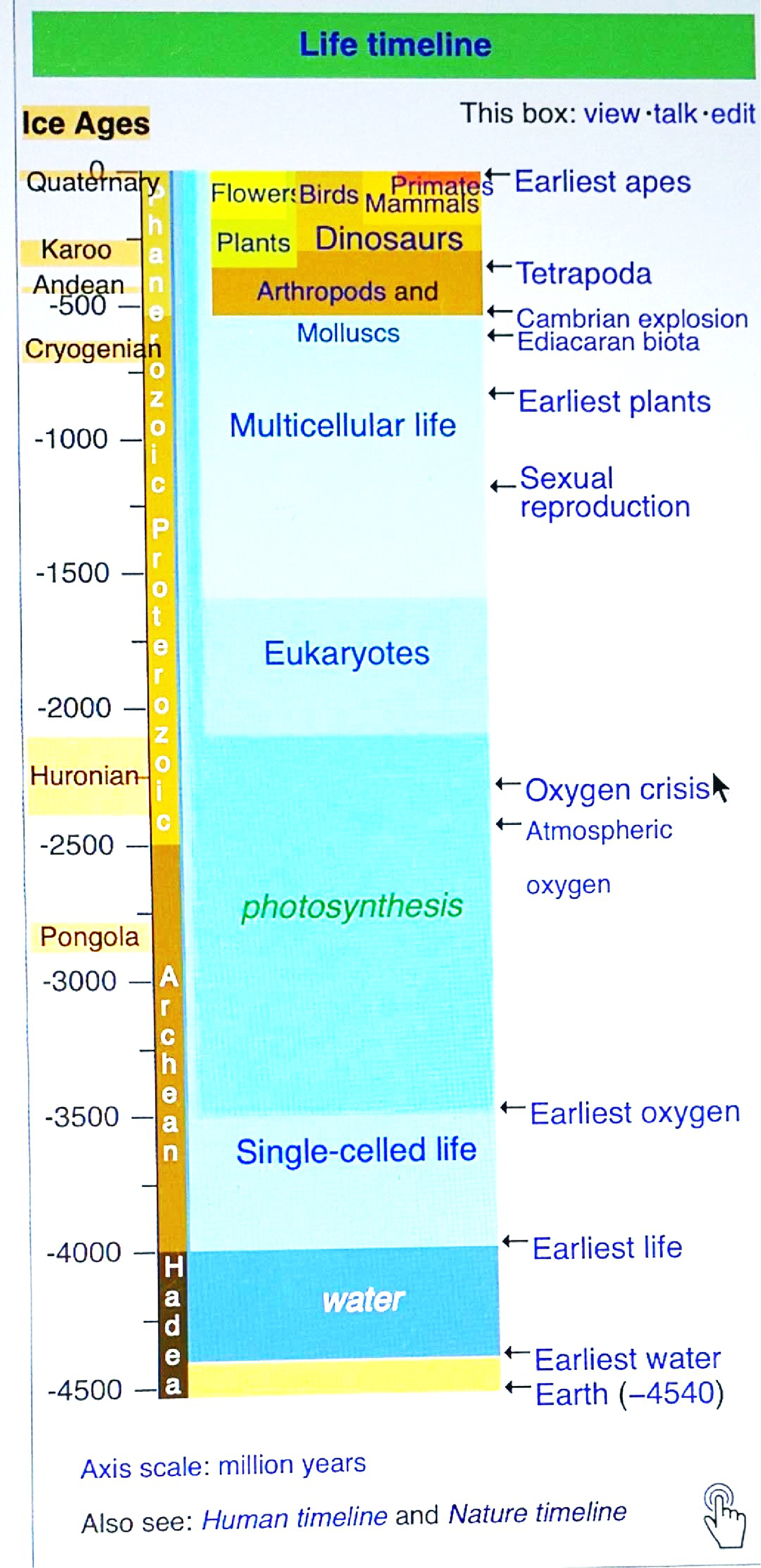
Photosynthetic organisms first appeared approximately 3.4-2.3 billion years ago
The carbon cycle in nature
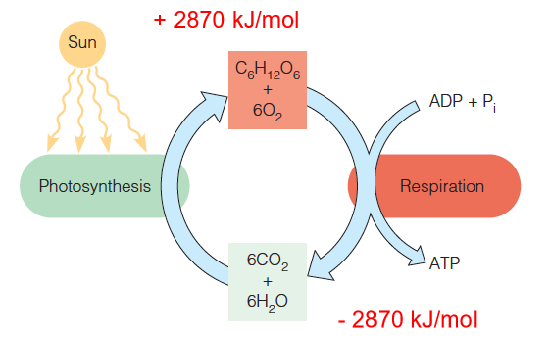
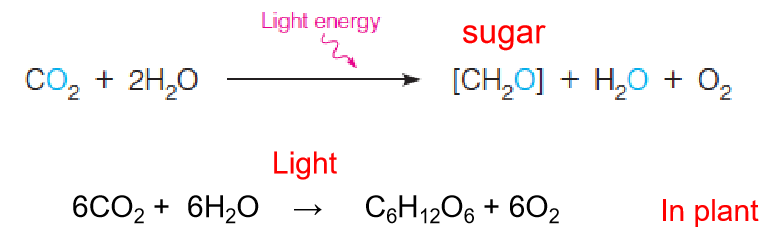
Carbon dioxide and water are combined through photosynthesis to form carbohydrates.
In both photosynthetic and nonphotosynthetic organisms these carbohydrates can be reoxidized to regenerate CO2 and H2O.
Part of the energy obtained from both photosynthesis and fuel oxidation is captured in ATP.
Photosynthesis requires a reductant, usually H2O (unlimited reducing equivalents), to reduce CO2 to the carbohydrate level.
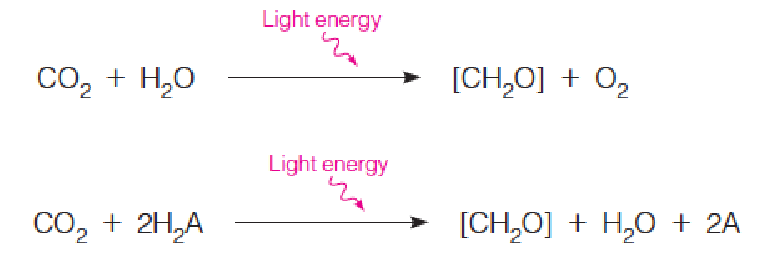
- [CH2O] represents a general carbohydrate
- CO2 is reduced to carbohydrate
- H2A: a general reductant
- A: oxidized product
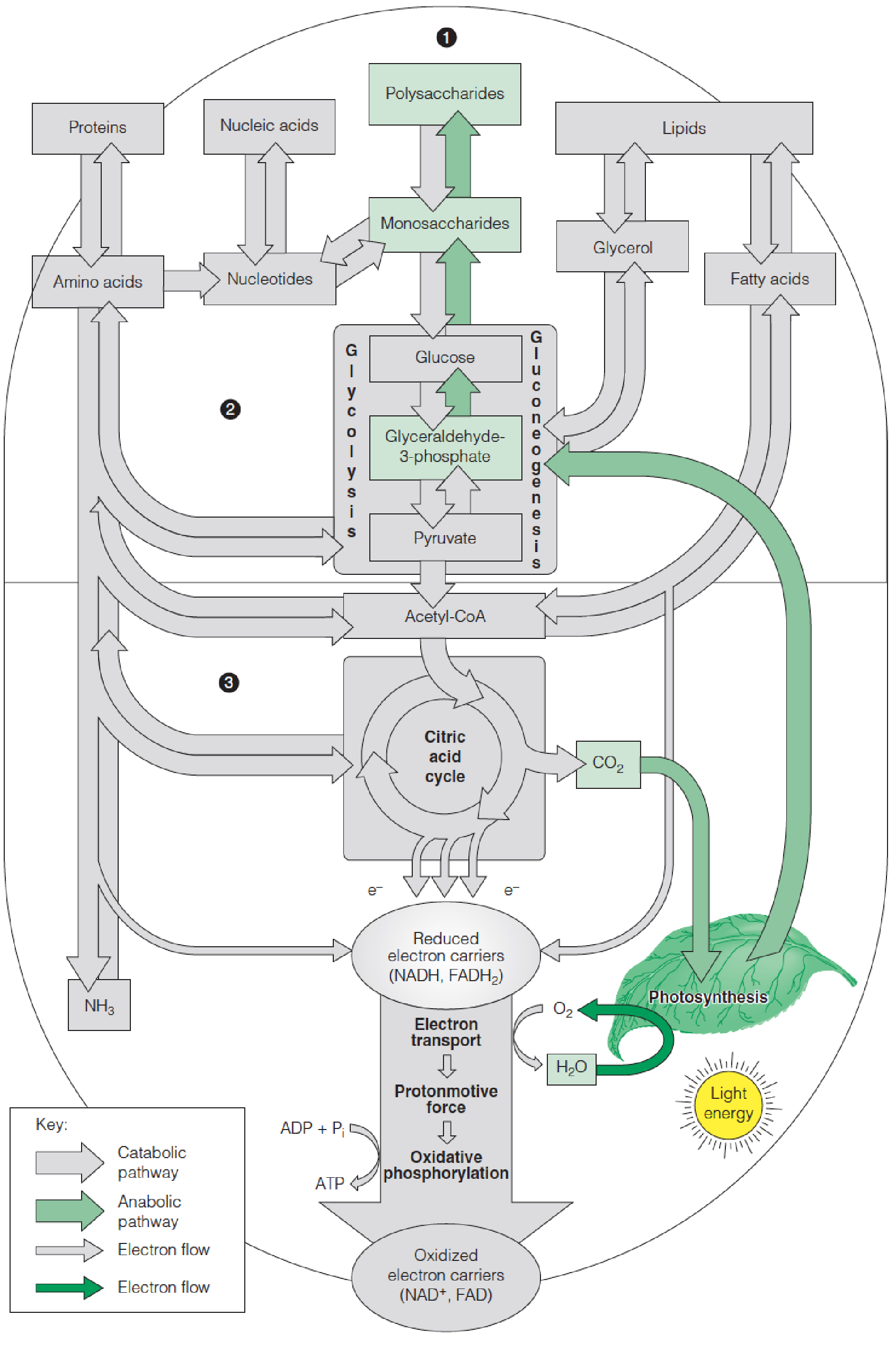
The role of photosynthesis in metabolism
The major biosynthetic pathways leading from carbon dioxide and water to polysaccharides are highlighted in green.
Oxygen derived from the water is released as a by-product of photosynthesis.
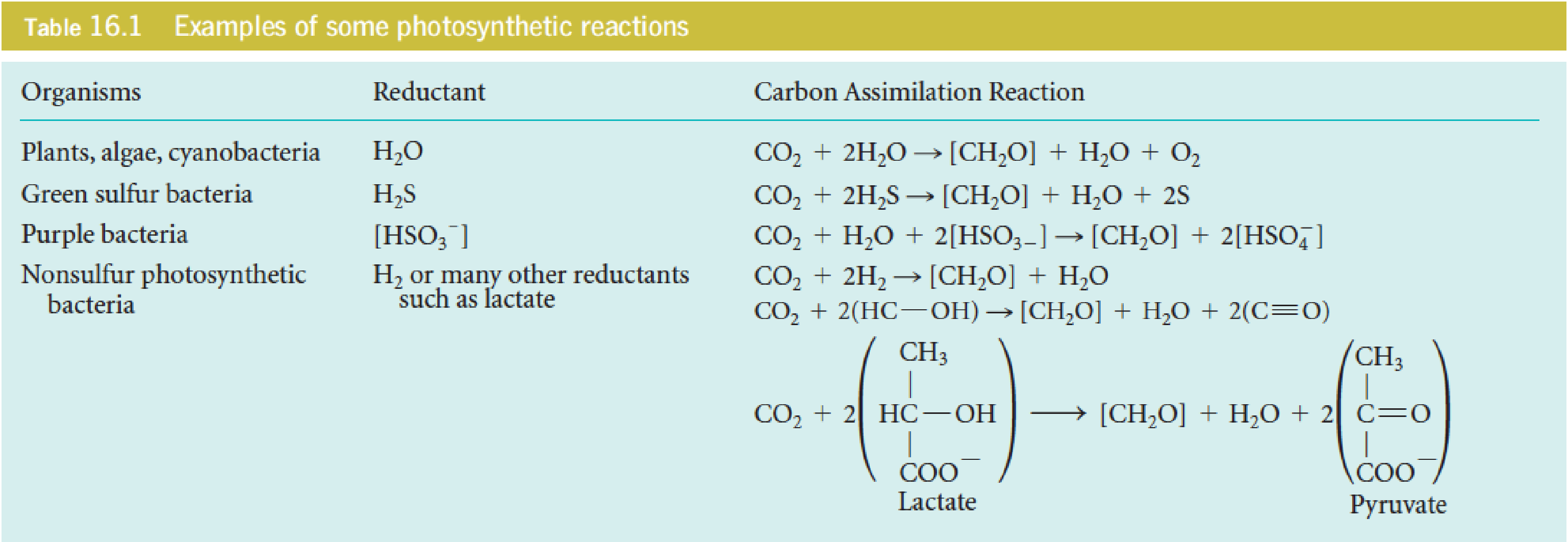
It is more correct to write the photosynthetic reactions in the fashion shown in Table 16.1, which makes it clear that one of the oxygens from CO2 ends up in carbohydrate, the other in water:
- ^18^O-labelled H2O, unlabelled CO2;
- Light energy cannot be used directly to drive this reaction;
- H2O does not reduce CO2 directly under any circumstances;
- The overall process is actually separated, both chemically and physically, into two subprocesses.
The two subprocesses of photosynthesis
The overall process of photosynthesis is divided into light reactions and dark reactions.
The light reactions, which require visible light as an energy source, produce reducing power (in the form of NADPH), ATP, and O2.
The NADPH and ATP drive the so-called dark reactions, which occur in both the presence and the absence of light and fix CO2 into carbohydrates.
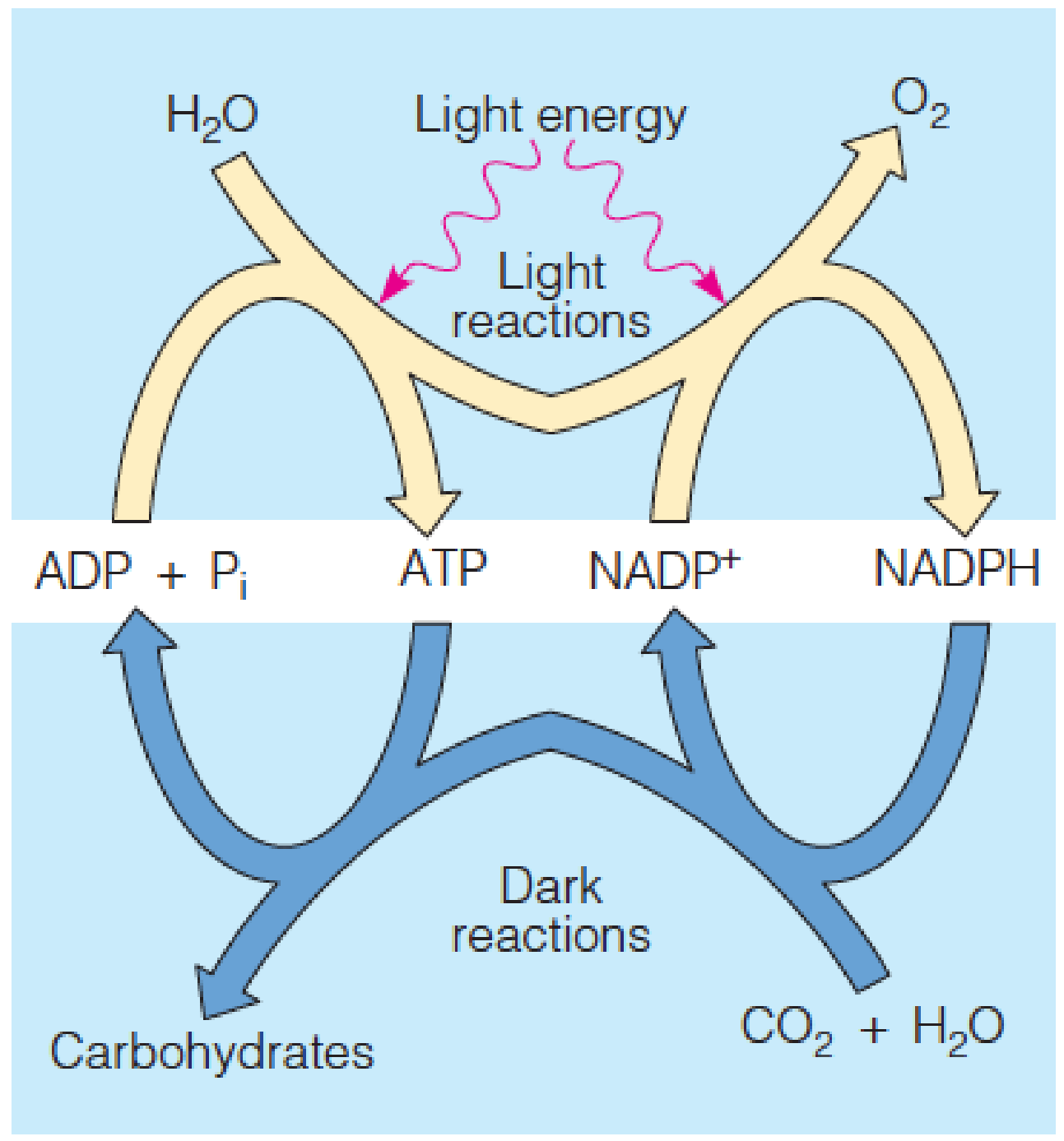
- H2O is the electron donor
Dark reactions do not require direct light participation; accelerated by light
二、The Chloroplast
Chloroplasts (叶绿体) are the photosynthetic organelles of green plants and algae.
Several chloroplasts are shown in a cross section of a cell from a Coleus(薄荷科植物) leaf.
Enlarged view of a single chloroplast from a leaf of timothy grass.(猫尾草)
Schematic rendering of a chloroplast.
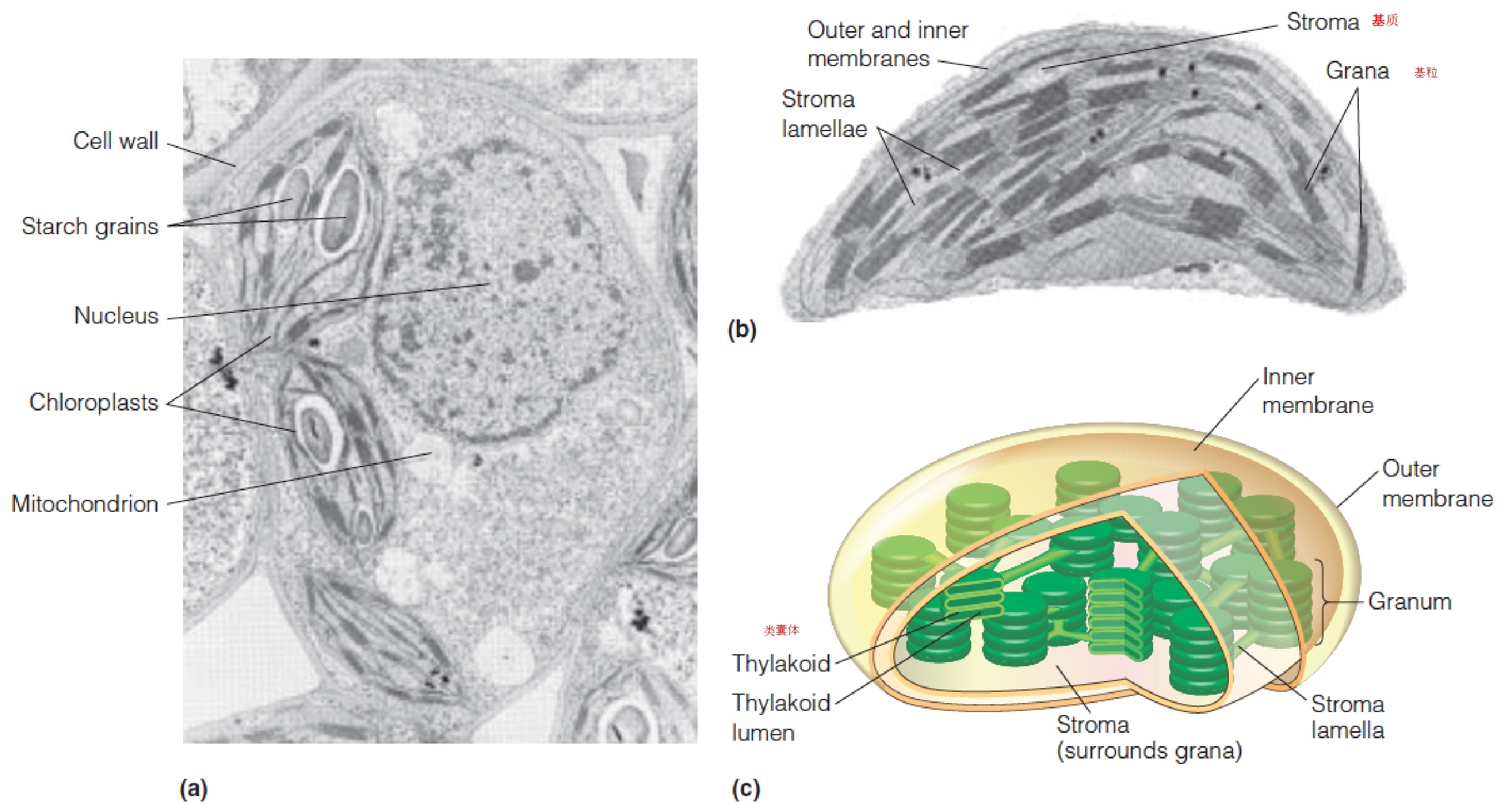
- Under the leaf surface (mesophyll cells(叶肉细胞))
- Each cell has 20-50 chloroplasts
- Semiautonomous, DNA, mRNA
- Inner and outer membrane, stroma, stroma lamella, thylakoid, grana, lumen
A photosynthetic prokaryote
This electron micrograph of a thin section of the cyanobacterium(藻青菌) Anabaena azollae(鱼腥藻) shows the folded membranes, which resemble the thylakoids(类囊体) of eukaryotic chloroplasts.
Absorption of light and the light reactions occur in the chloroplast membranes.
The dark reactions occur in the stroma.
三、The Light Reactions
The energy of photons
The graph shows energy per mole of photons as a function of wavelength, compared with energies of several chemical bonds.
Light in the ultraviolet range has enough energy to break many chemical bonds directly.
Visible light can break some weak bonds.
Light in the long-wavelength portion of infrared region of the spectrum causes only heat-producing molecular vibrations.
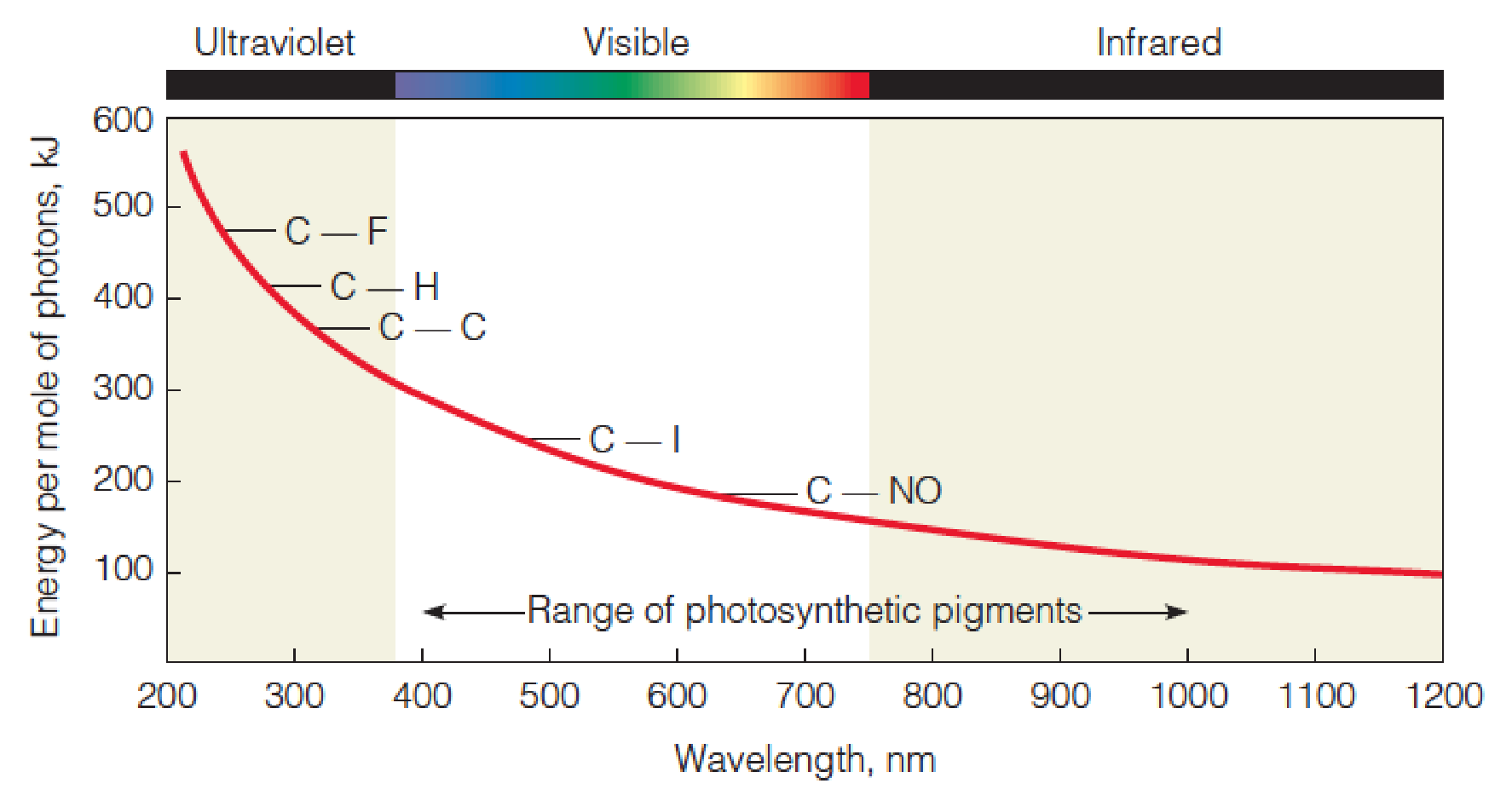
- Ozone layer blocks 97% far-ultraviolet; ozone depletion
- Photosynthesis: visible + near- infrared (400-1000)
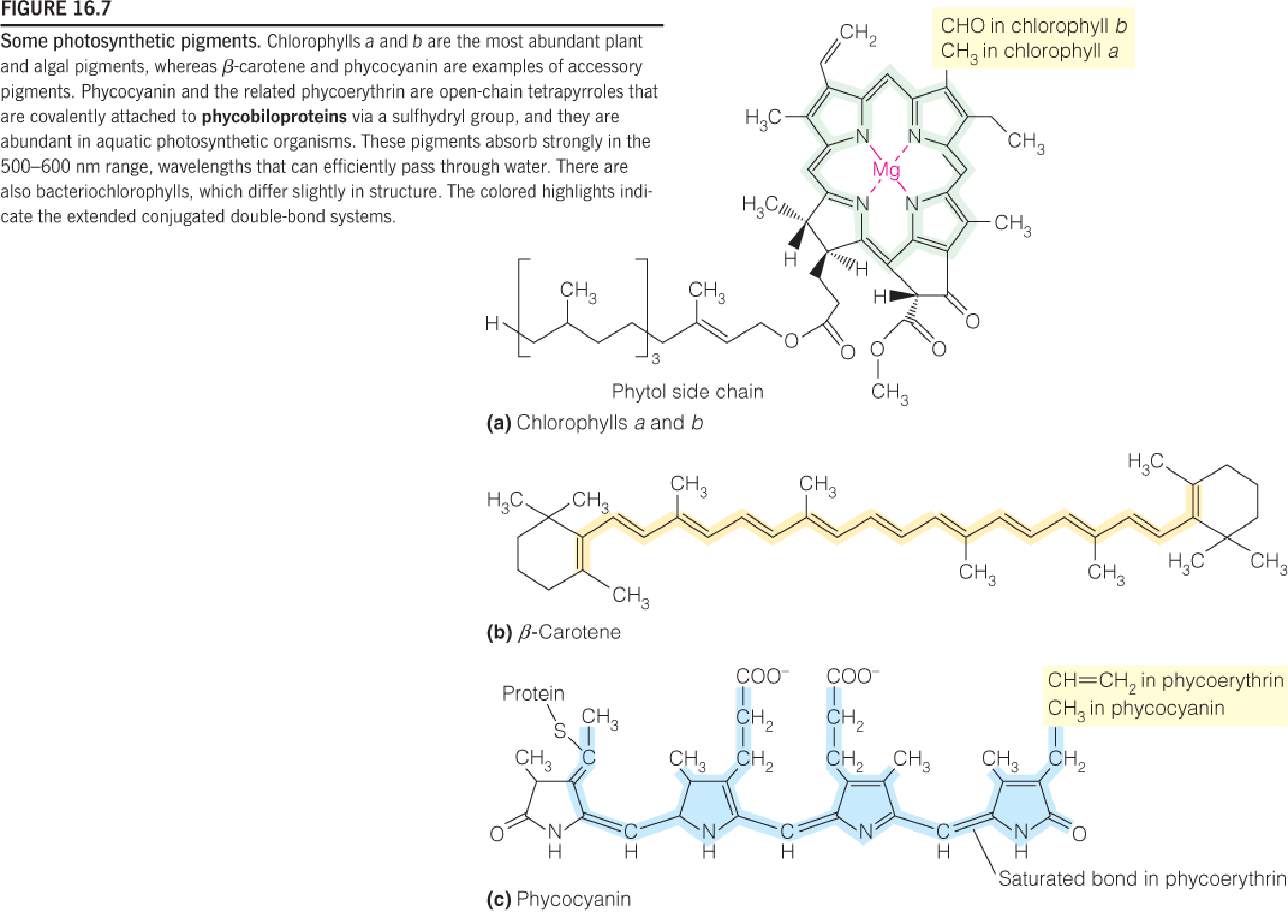
Some photosynthetic pigments
Chlorophylls(叶绿素) a and b are the most abundant plant and algal pigments, whereas β-carotene and phycocyanin(藻蓝蛋白) are examples of accessory pigments.
Phycocyanin and the related phycoerythrin(藻红蛋白) are open-chain tetrapyrroles that are covalently attached to phycobiloproteins(藻胆蛋白) via a sulfhydryl group, and they are abundant in aquatic photosynthetic organisms.
These pigments absorb strongly in the 500–600 nm range, wavelengths that can efficiently pass through water.
There are also bacteriochlorophylls, which differ slightly in structure.
- Absorption spectra and light energy
The absorption spectra of various plant pigments are compared with the spectral distribution of the sunlight that reaches the earth’s surface.
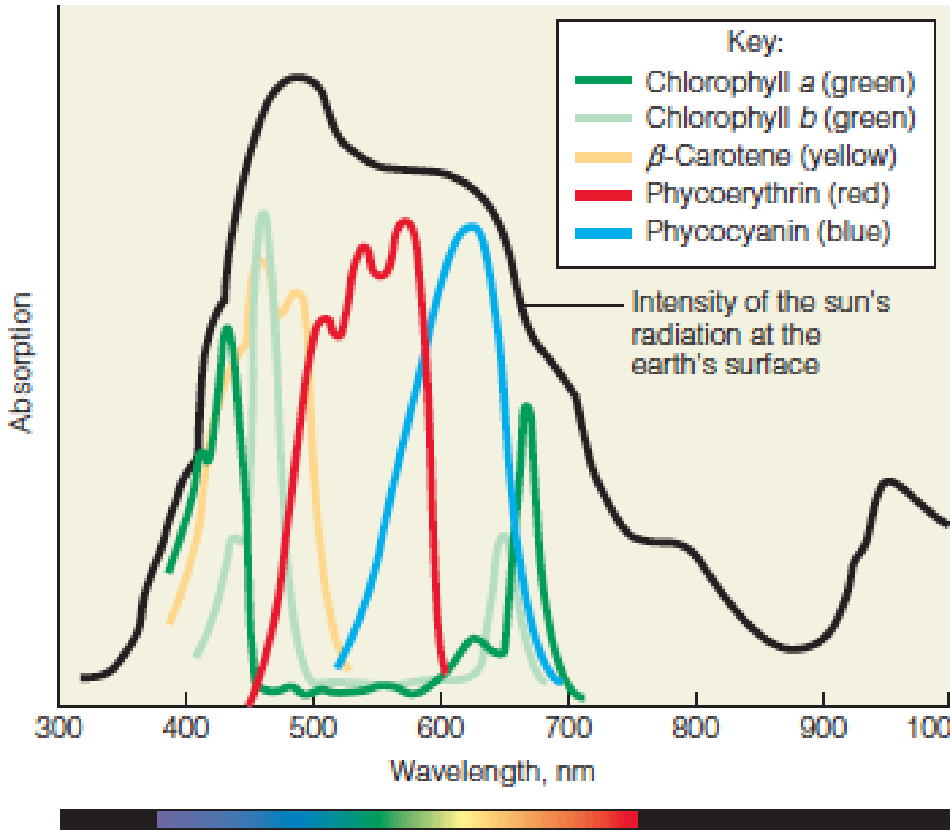
- Chlorophyll a and b absorb strongly in both the deep blue and red, the light that is NOT absorbed but reflected from chloroplasts is green. The color of most growing plants;
- The other observed colors, red, brown or purple—accessary pigments;
- Loss of a and b in autumn leaves allows accessary pigments and nonphotosynthetic pigments to become evident, such as β-carotene.
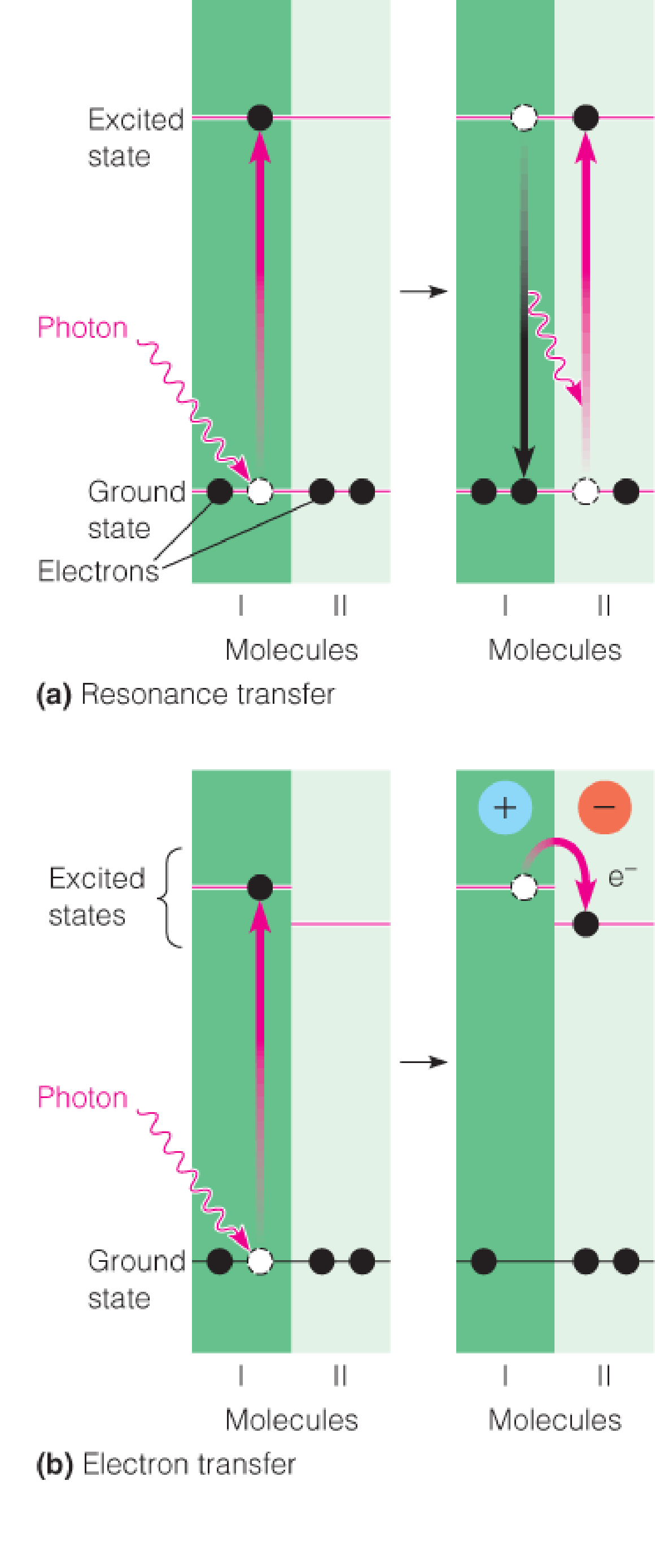
Two modes of energy transfer following photoexcitation
- In resonance transfer(共振转移) molecule I transfers its excitation energy to an identical molecule II, which rises to its higher energy state as molecule I falls back to the ground state. Resonance transfer is extraordinarily fast.
- In electron transfer an excited electron in molecule I is transferred to the slightly lower excited state of molecule II, making molecule I a cation and molecule II an anion.
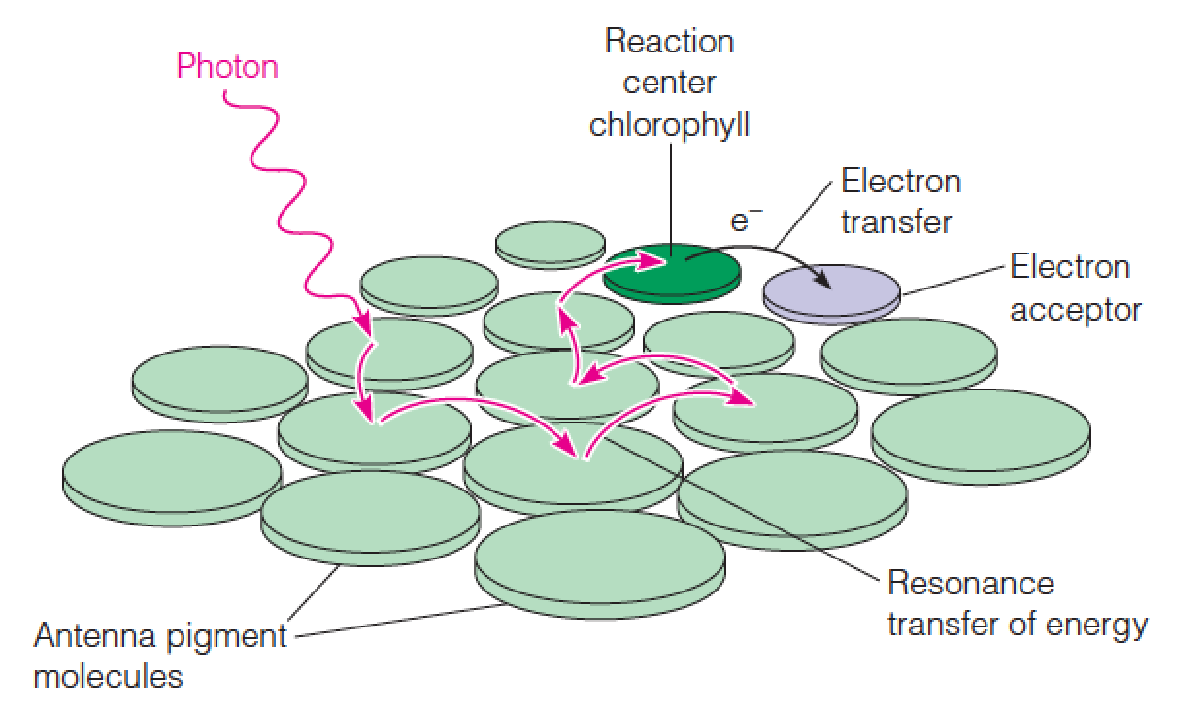
Most chlorophyll molecules are used as antennae(天线) to catch photons and pass their energy on to reaction centers.
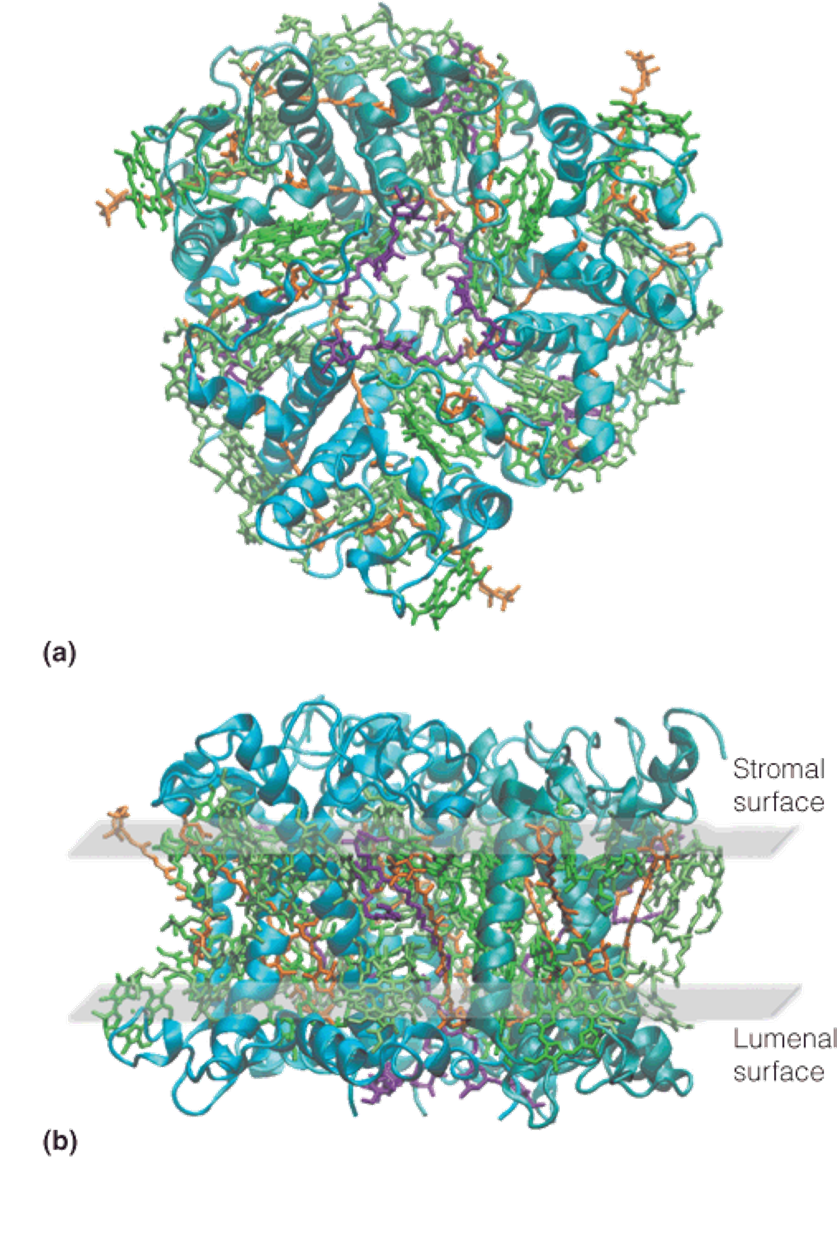
Three-dimensional structure of the trimeric light-harvesting complex II of plants.
The X-ray structure of the pea LHCII shows that the protein exists as a homotrimer buried in the thylakoid membrane.
Each trimer contains 24 chlorophyll a molecules, 18 chlorophyll b molecules, and 12 carotenoids(类胡萝卜素) (luteins and xanthophylls), all serving as antenna molecules.
1. Resonance transfer of energy in a light-harvesting complex:
The excitation energy originating in a photon of light wanders(徘徊,漫步) from one antenna molecule to another until it reaches a reaction center.
There an electron is transferred to a primary electron acceptor molecule, and the energy is trapped.
Two photosystems, linked in series, are involved in the photosynthetic light reactions in algae, cyanobacteria, and higher plants.
In each of the two photosystems, the primary step is transfer of a light-excited electron from a reaction center (P680 or P700) into an electron transport chain.
The ultimate source of the electrons is the water molecules producing O2.
The final destination of the electrons is the molecule of NADP^+^, which is thereby reduced to NADPH.
At two stages in the electron transport process, protons are released into the thylakoid (类囊体) lumen.
Some of the protons come from the H2O that is broken down, some come from the stroma.
This transfer of protons into the lumen produces a pH gradient across the thylakoid membrane.
Thus, ATP and reducing power in the form of NADPH (reducing equivalent) are the products of the light reactions.
These compounds are exactly what is needed to drive the syntheses carried out in the dark reactions.
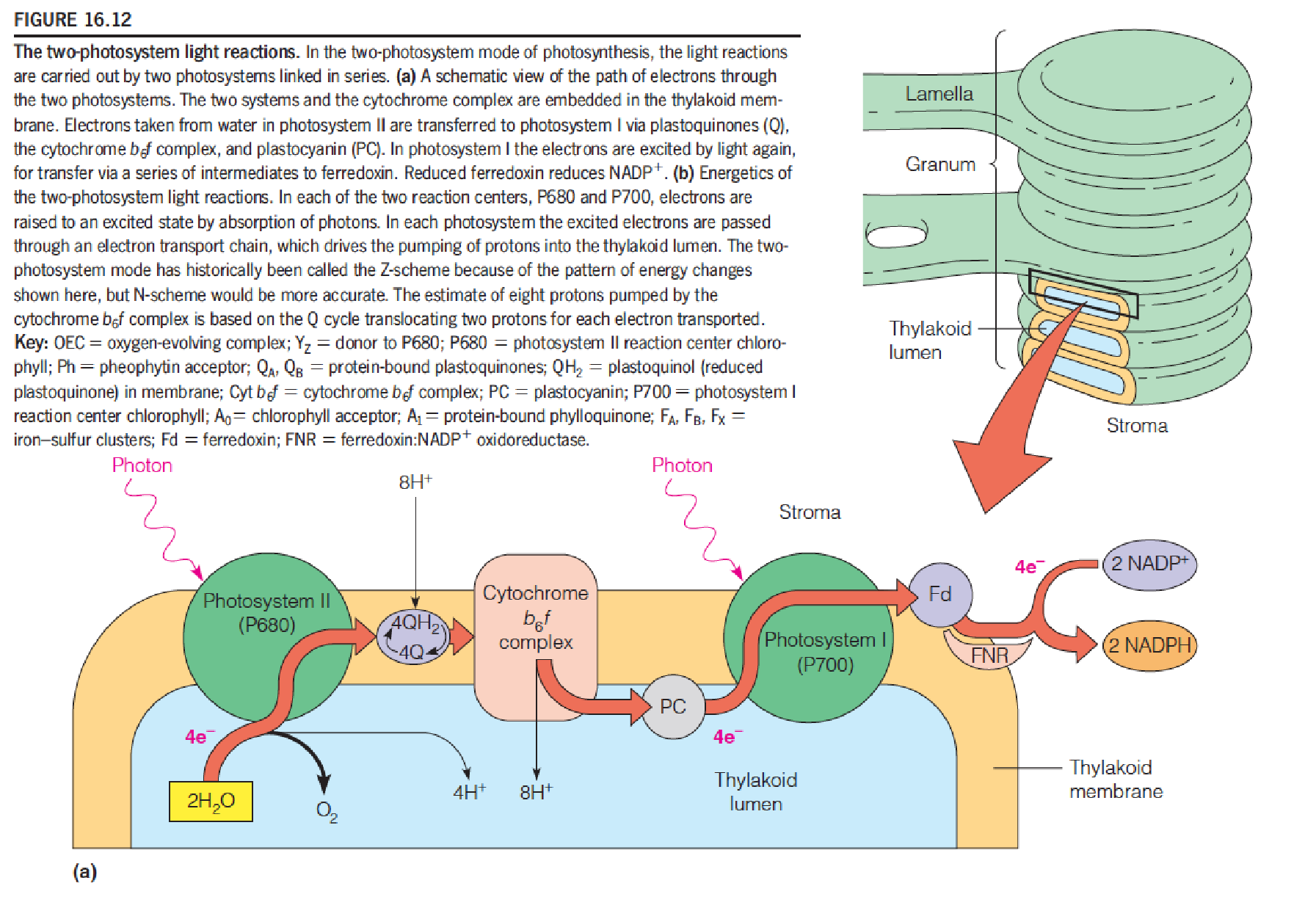
- Photosystem Ⅱ: P680
- Photosytem Ⅰ: P700
- H+的两个来源:(1) H
2O (2) pumped by Cytochrome b6f
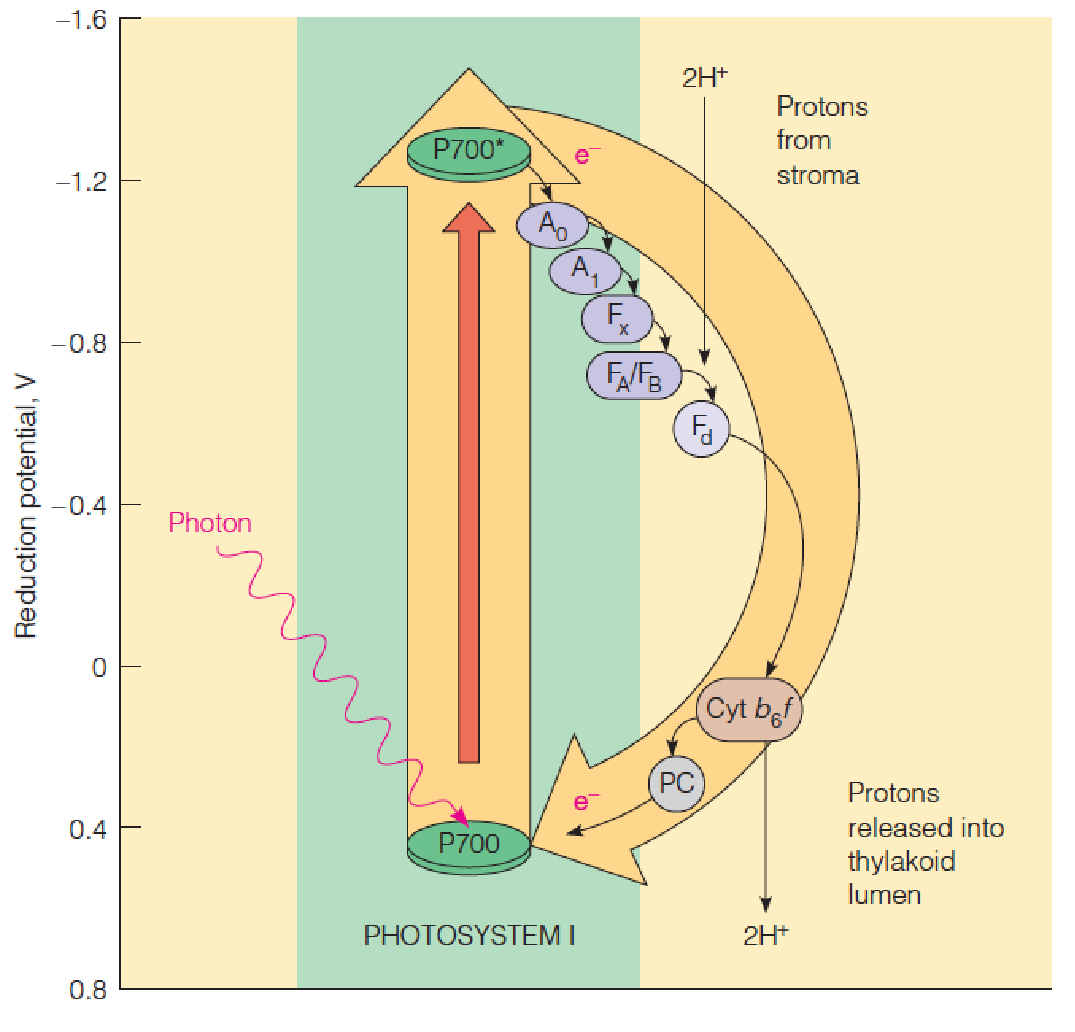
2. Energetics 动力学 of the two-photosystem light reactions
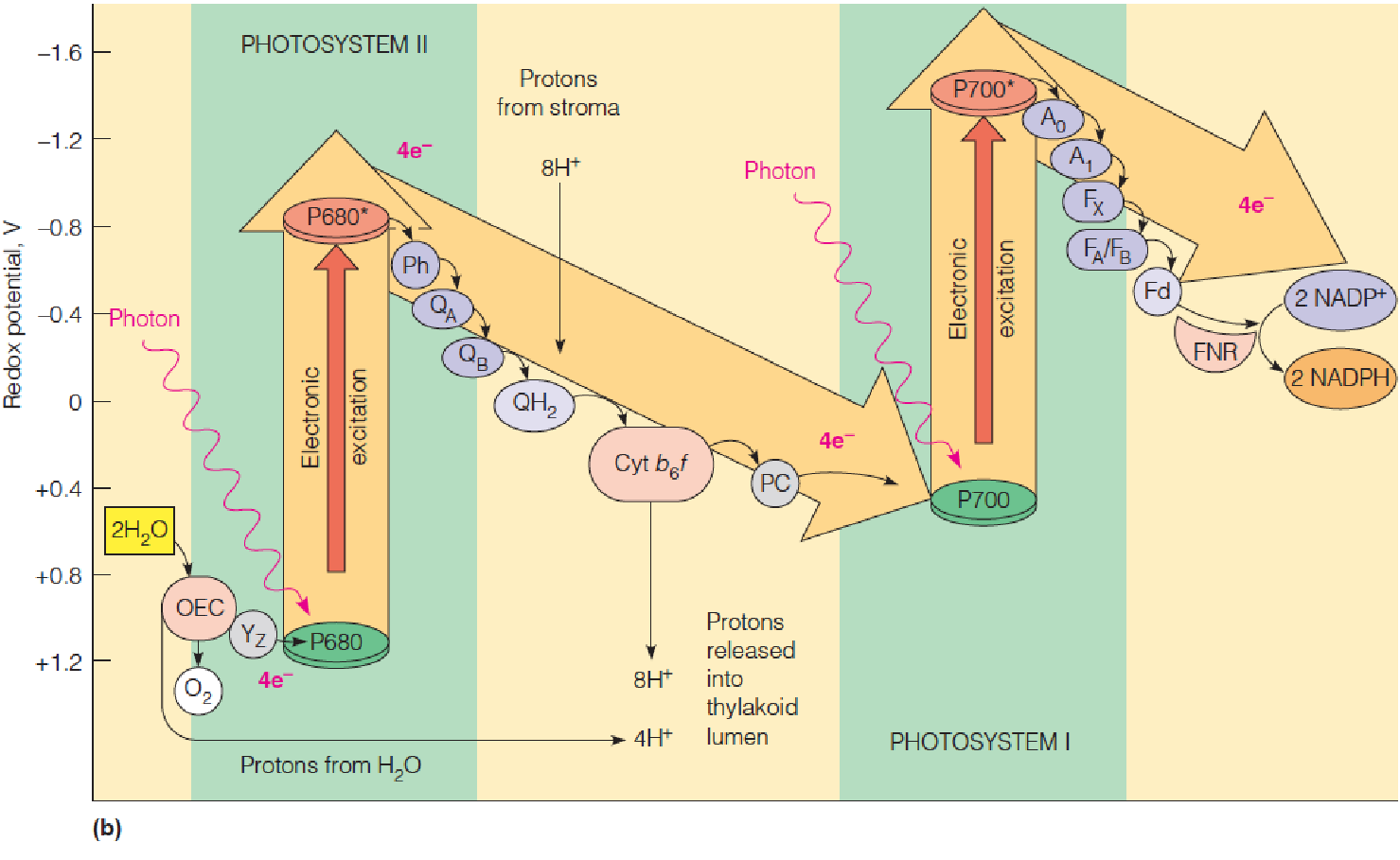
- OEC: oxygen-evolving complex;
- Yz: donor to P680; P680:
- PSII reaction center chlorophyll;
- Ph: phyeophytin acceptor;
- QA,QB: protein-bound plastoquinones;
- QH2: plastoquinol (reduced plastoquinone) in membrane;
- cyt b6f: cytochrome b6f complex;
- PC: plastocyanin;
- P700: PSI reaction center chlorophyll;
- A0: chlorophyll acceptor;
- A1: protein-bound phylloquinones;
- FA, FB, FX: ion-sulfur clusters;
- Fd: ferredoxin;
- FNR: ferredoxin:NADP+ oxidoreductase
Electrons taken from water in PSII are transferred to PSI via plastoquinone (Q), cyt b6f complex, and plastocyanin (PC, 质体蓝素). In PSI, electrons are excited again by photon (light) for transfer via a series of intermediates to ferredoxin and reduced ferredoxin reduces NADP+ to NADPH.
PC (plastocyanin, 质体蓝素):
- mobile protein (allowing long distance electrons transfer;
- pass e- to P700 reaction center.
2. Structure and reactivity of plastoquinone(质体醌)
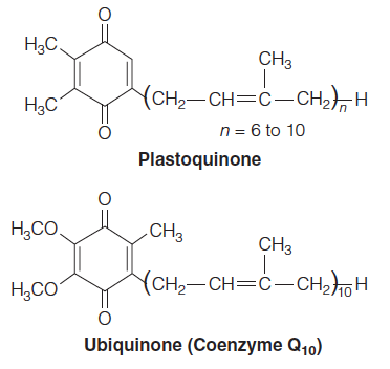
Plastoquinone(质体醌) is similar to ubiquinone (CoQ,泛醌)
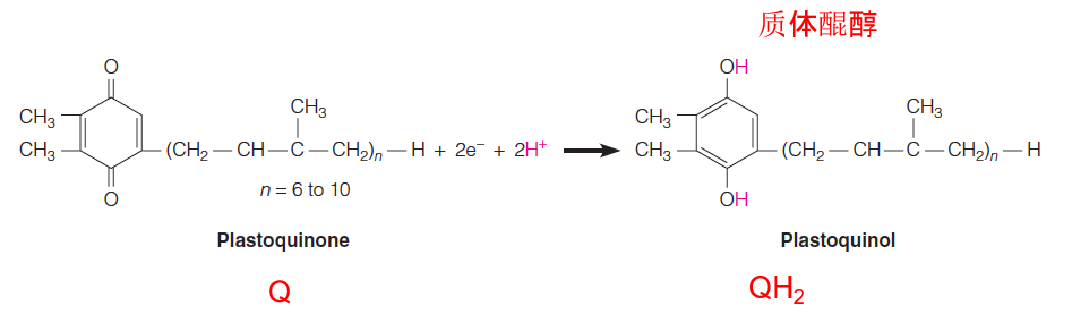
- Q和CoQ具有相同的functional group
3. Structure of the cytochrome b6f complex
b6f:
- transfer e- to PSI;
- pump protons from stroma to thylakoid lumen (2H+ per electron)
- Similar to complex III
- Similar Q cycle
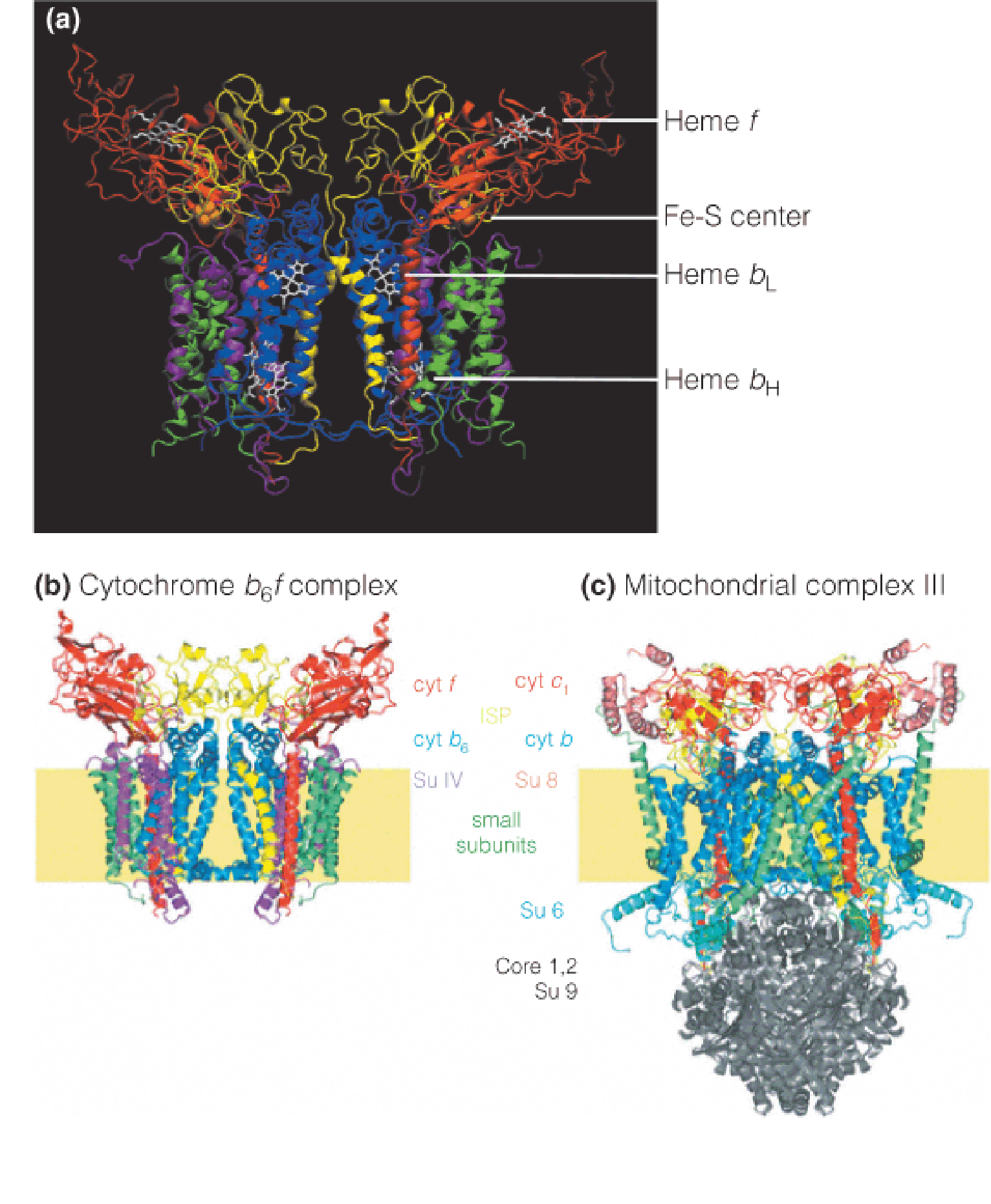
4. Structure of photosystem II
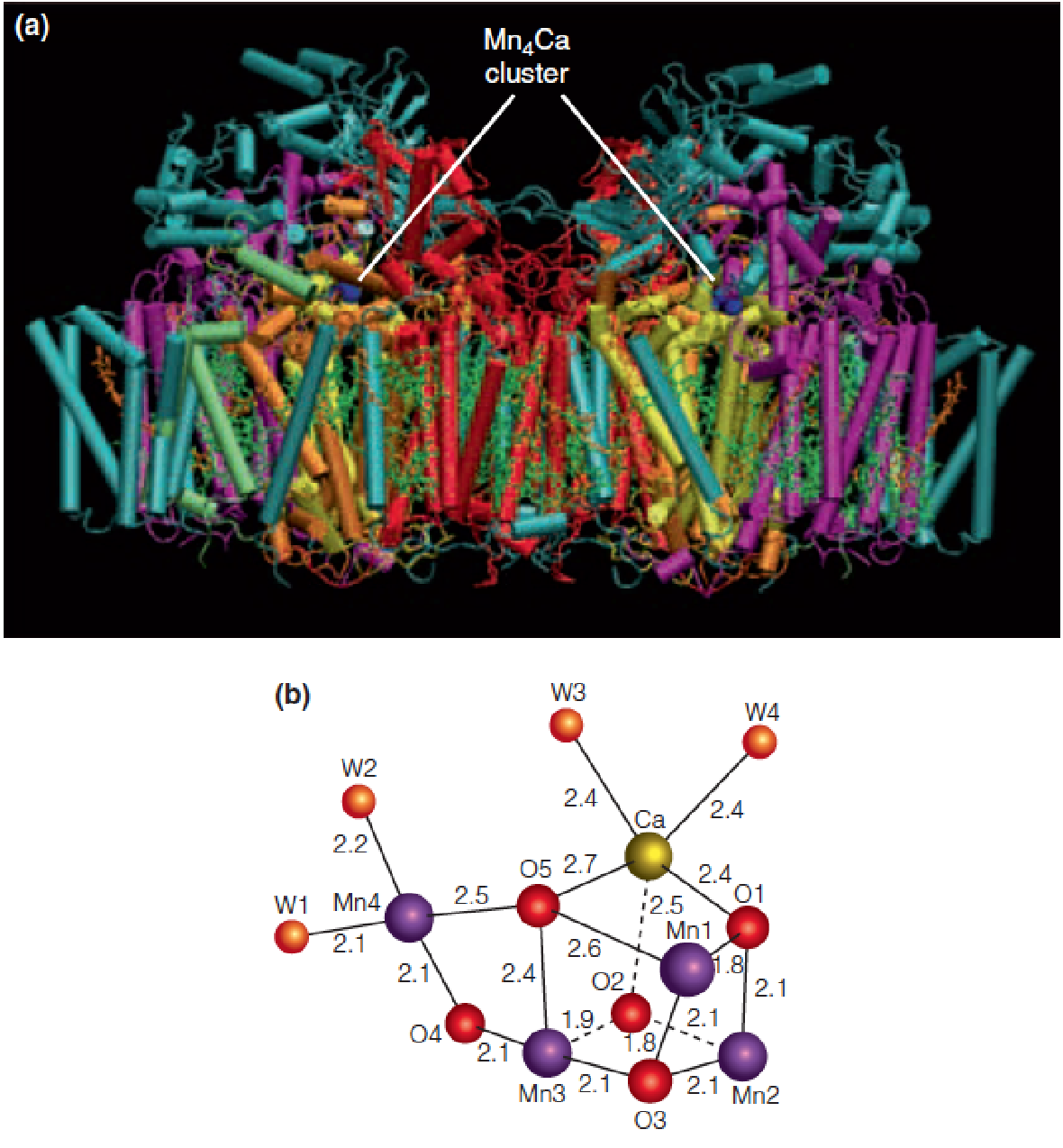
- Thylakoid membrane 位于类囊体的膜上
- multiple subunits, each monomer: 20 subunits (D1, D2)
- Water splitting catalytic component
- OEC: Mn
4Ca
- OEC: Mn
- P680 → P680^+^ + e-
5. The structure of plant photosystem I
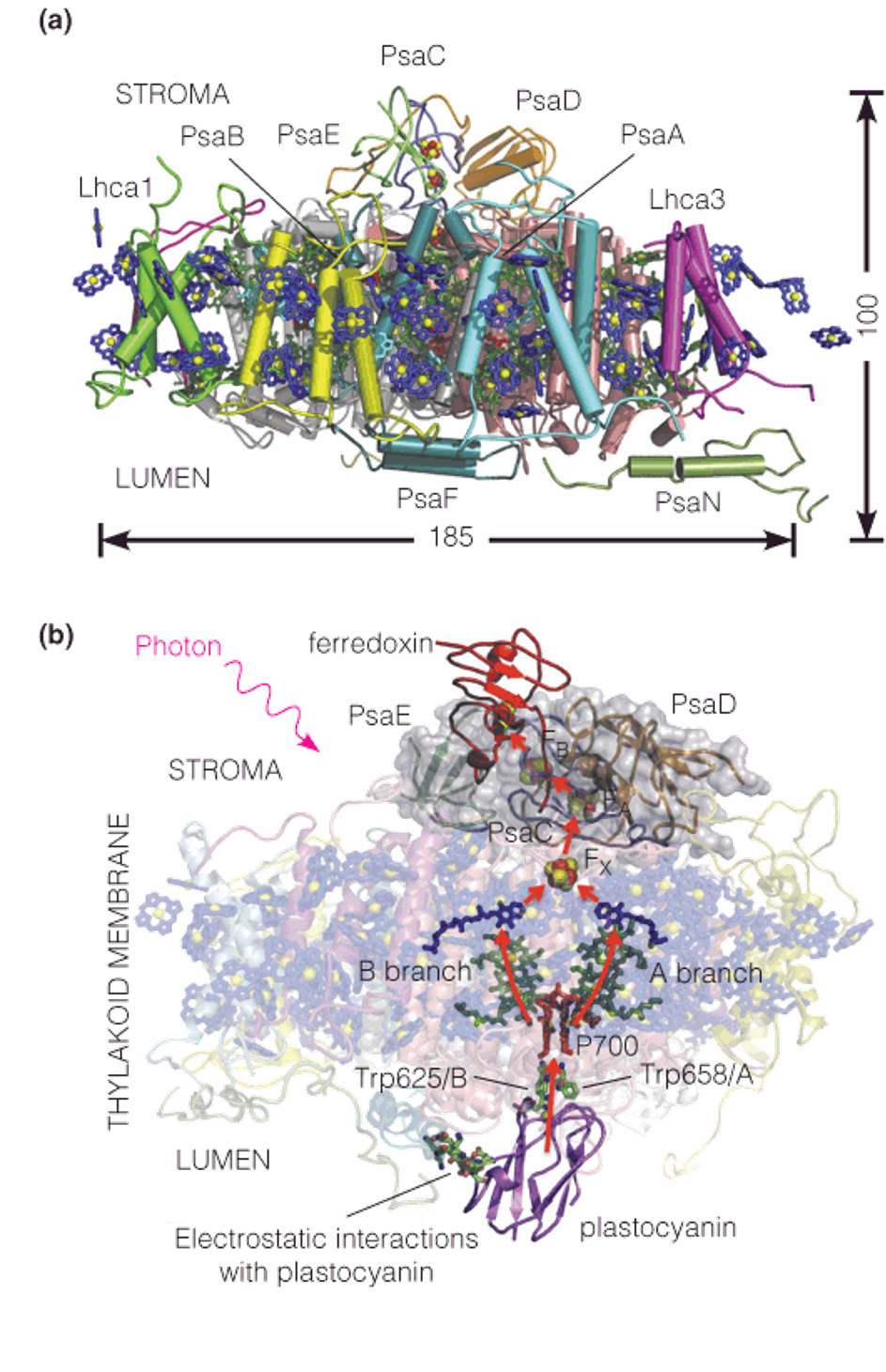
- 19 proteins
- Many antenna chlorophylls
- A reaction center chlorophyll, P700, which absorbs light of up to 700 nm
- A special chlorophyll electron acceptors (A0) → A1 → Fx → FA → Fd → NADP+ → NADPH
- Fd (ferredoxin): direct recipient of electrons
A model for the catalytic function of the oxygen-evolving complex (OEC) cluster in PSII
S0 - S4 represent the different oxidation states that the ligated metal cluster cycles through as e- and H+ are abstracted from the H2O molecules.
In the first four transitions, light energy is used to oxidize P680 to P680+, which in turn oxidizes the metal-oxo cluster of the OEC.
The S0 → S4 transition is light-independent and releases O2. Thus, four photons are required to oxidize two H2O to one O2.
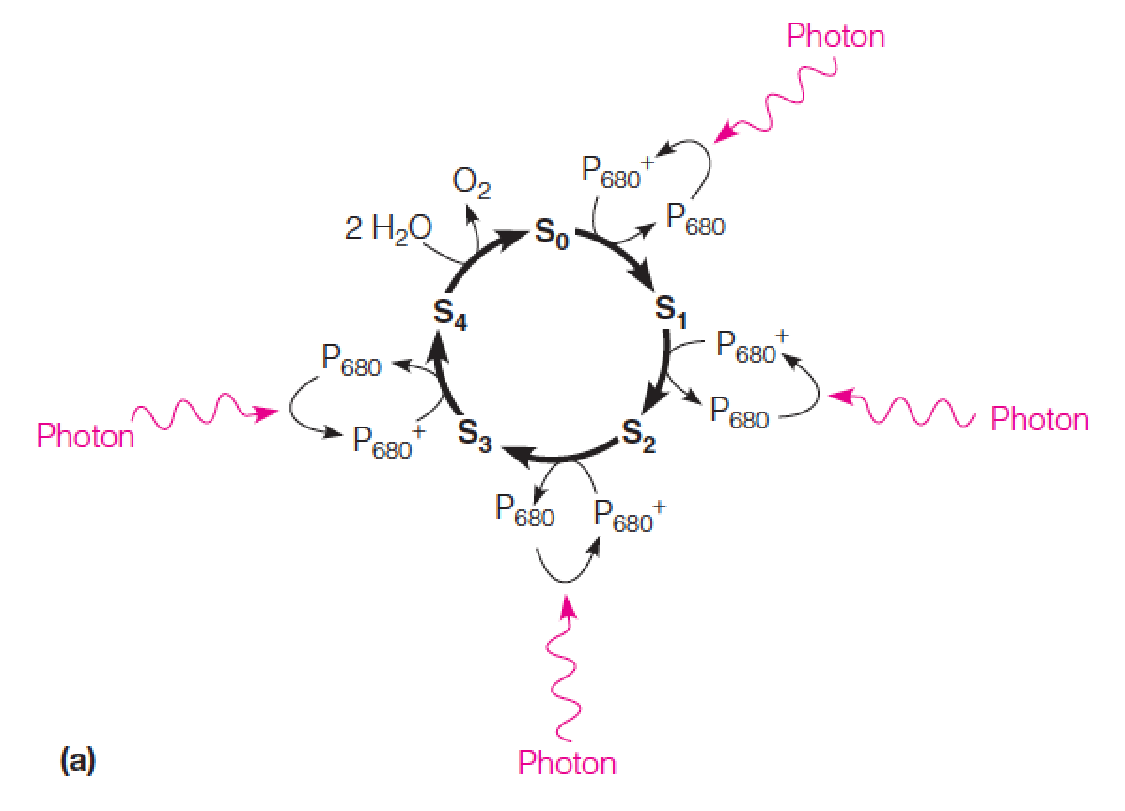
The Splitting of Water by PSII
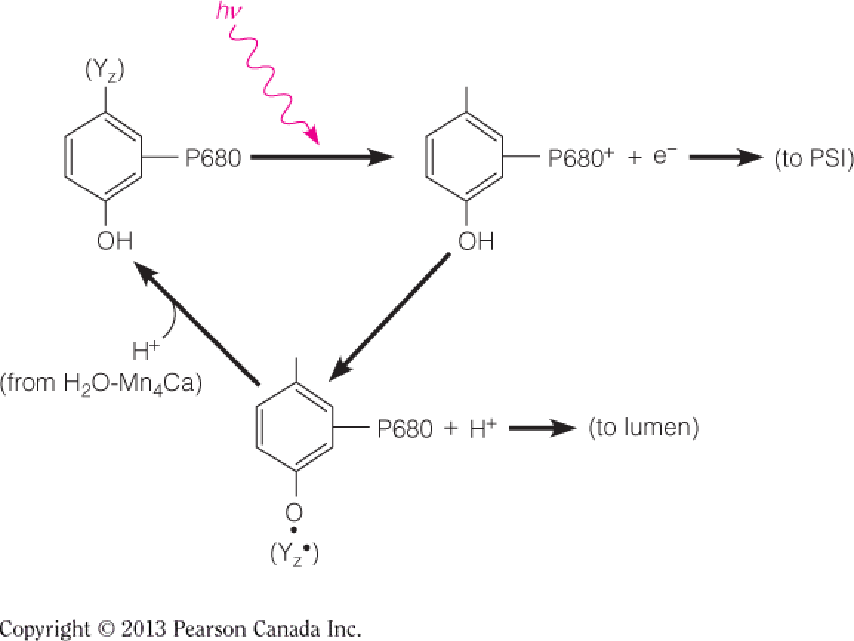
Detailed Process
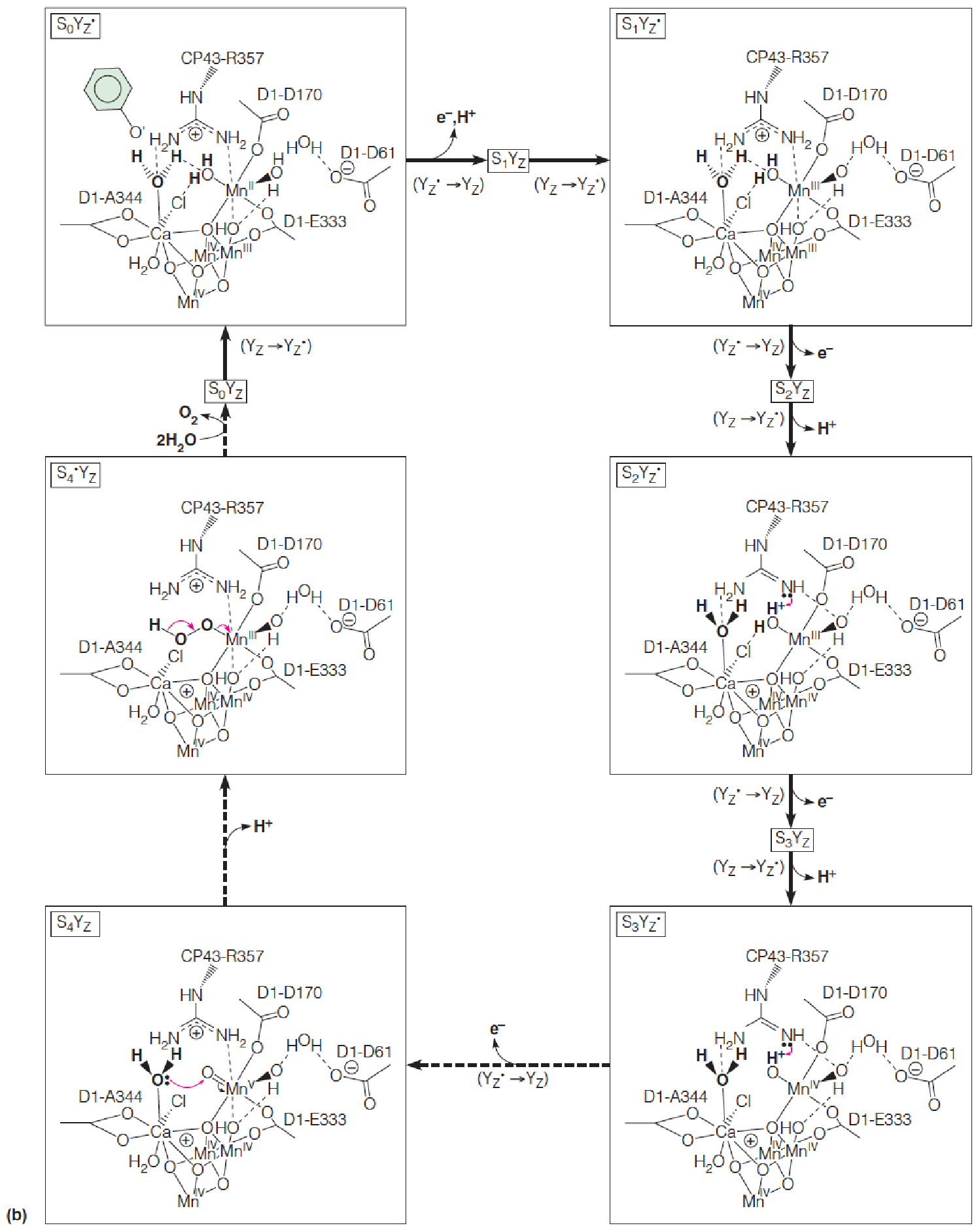
In this proposed mechanism, YZ represents the redox-active tyrosine residue of the D1 subunit; the oxidized form of YZ is the neutral, deprotonated radical species, YZ.
The two H2O molecules are bound in the S0YZ state by the Ca2+ ion and the “dangler” Mn^III^ ion.
Each transition passes through an intermediate SnYZ , in which the oxidized tyrosine is poised to oxidize the metal cluster of the OEC, but for clarity, the tyrosine radical is shown only once.
In this model, only two of the Mn ions change their oxidation states.
The S4→S4’ transition is the crucial O-O bond-forming step and involves the attack by a calcium-bound water upon the electrophilic Mn^V^=O oxo group. O2 is released, and two new H2O are bound to initiate another cycle.
- Mn价态的改变最终使得H2O splitting
Simple Summary
The system has in effect stripped four electrons from the four hydrogen atoms in two water molecules.
The oxygen produced diffuses out of the chloroplast.
The four protons that are produced from the two water molecules are released into the thylakoid lumen, helping to generate a pH difference between the lumen and stroma.
We may summarize the reaction carried out by photosystem II as follows:
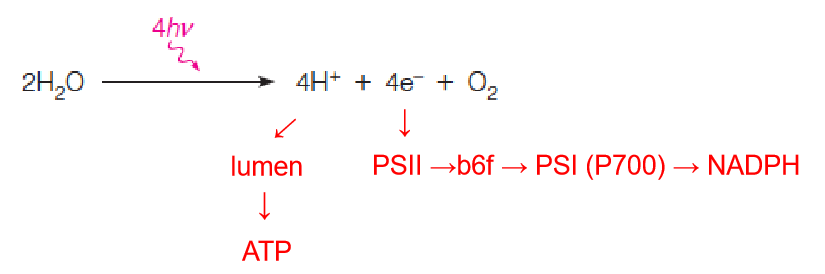
Summary view of the light reactions as they occur in the thylakoid
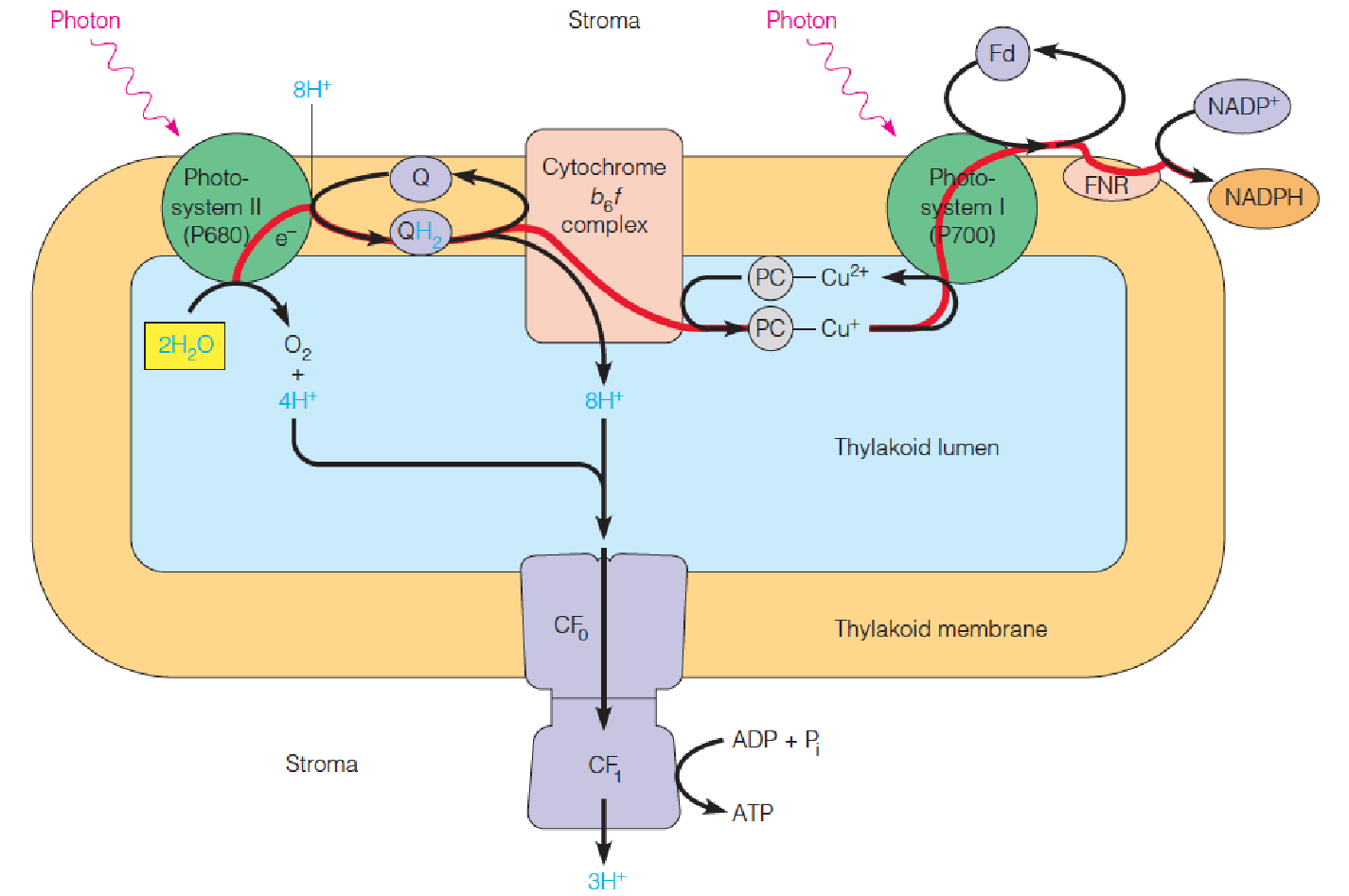
Arrangement of components of the two photosystems on the thylakoid membrane
- The membrane layers in the interior of the granum(基粒) are rich in photosystem II.
- The stroma lamellae(基质片层) and the top and bottom surfaces of the granum are rich in photosystem I and ATP synthase particles, allowing NADP reduction and ATP generation to occur at or near these stroma-facing surfaces.
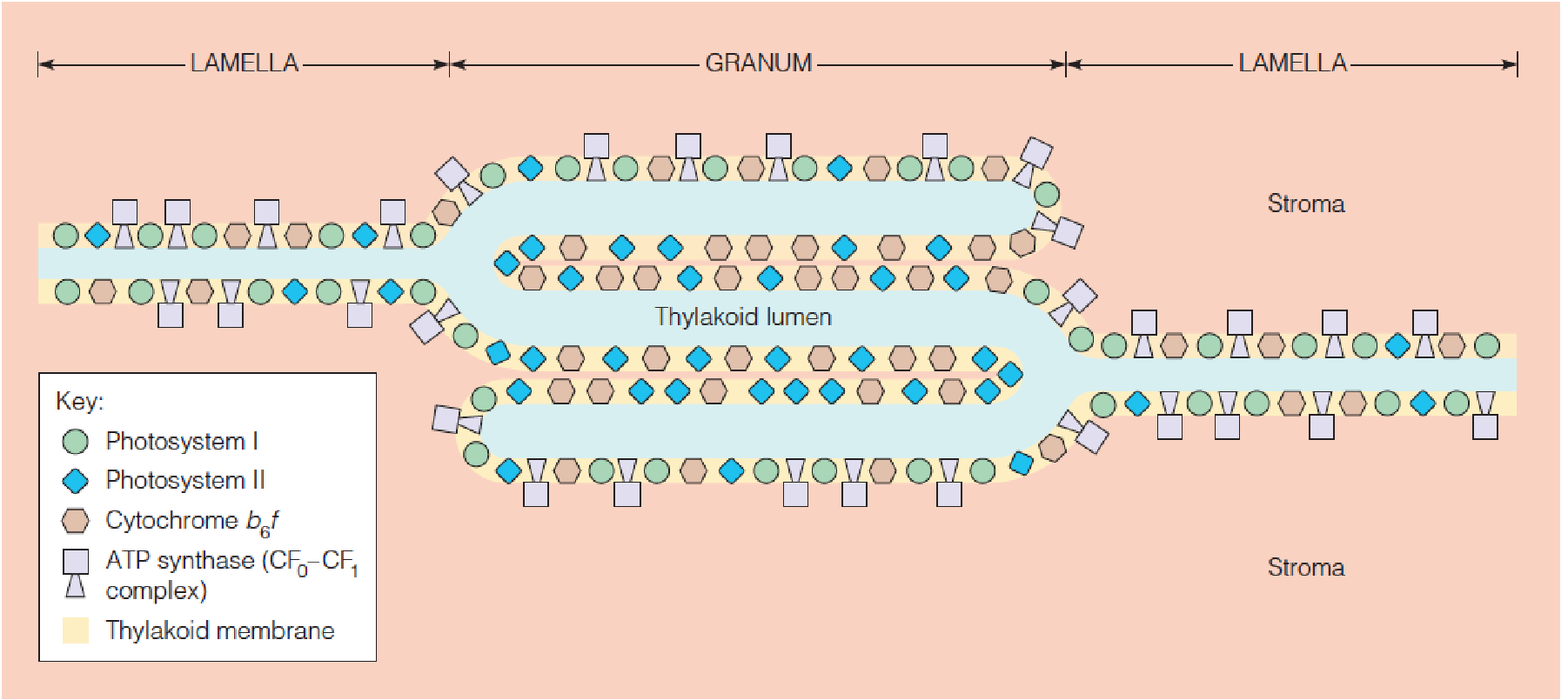
- CF0-CF1 complex利用lumen内部的H+ gradient合成ATP到stroma,它与F0F1 complex结构具有一定的相似性
Cyclic electron flow 循环电子流
When levels of NADP+ are low and levels of NADPH are high, electrons from the P700 center are returned to it via the cytochrome b6f complex.
There is no NADP+ reduction, but protons are pumped across the membrane and therefore ATP is generated.
Only in photosystem I:
- An alternative light reaction mechanism; 电子回到b6f,pump proton from stroma to lumen,形成了H+ gradient,因此没有NADPH的合成
- cyclic photophosphorylation (ATP);
- no NADPH production;
- no O2 is released;
- Helps maintain balance between ATP and NADPH production.
Model of a purple bacterial reaction center complex
Photosynthetic bacteria:
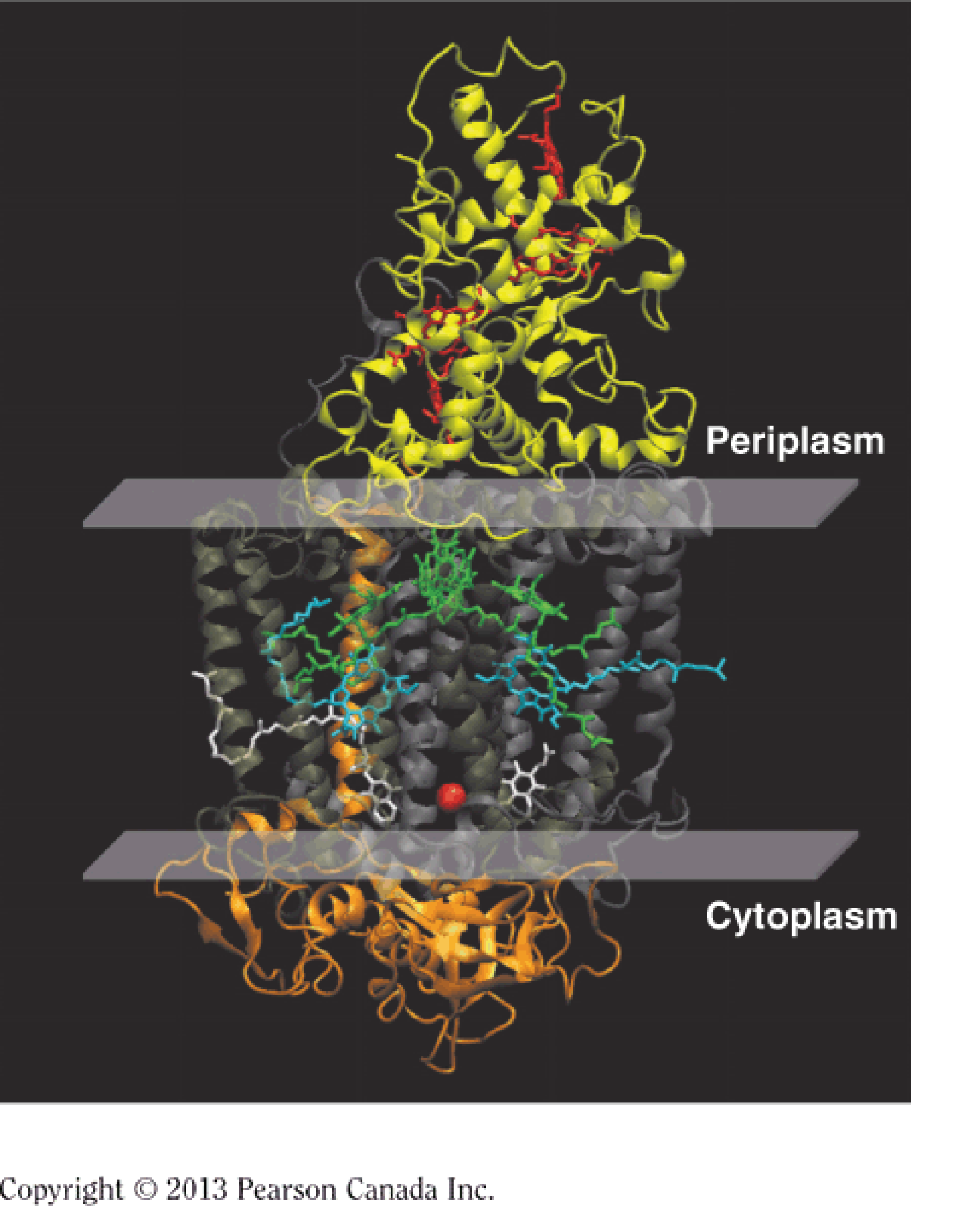
Nobel Prize in Chemistry in 1988
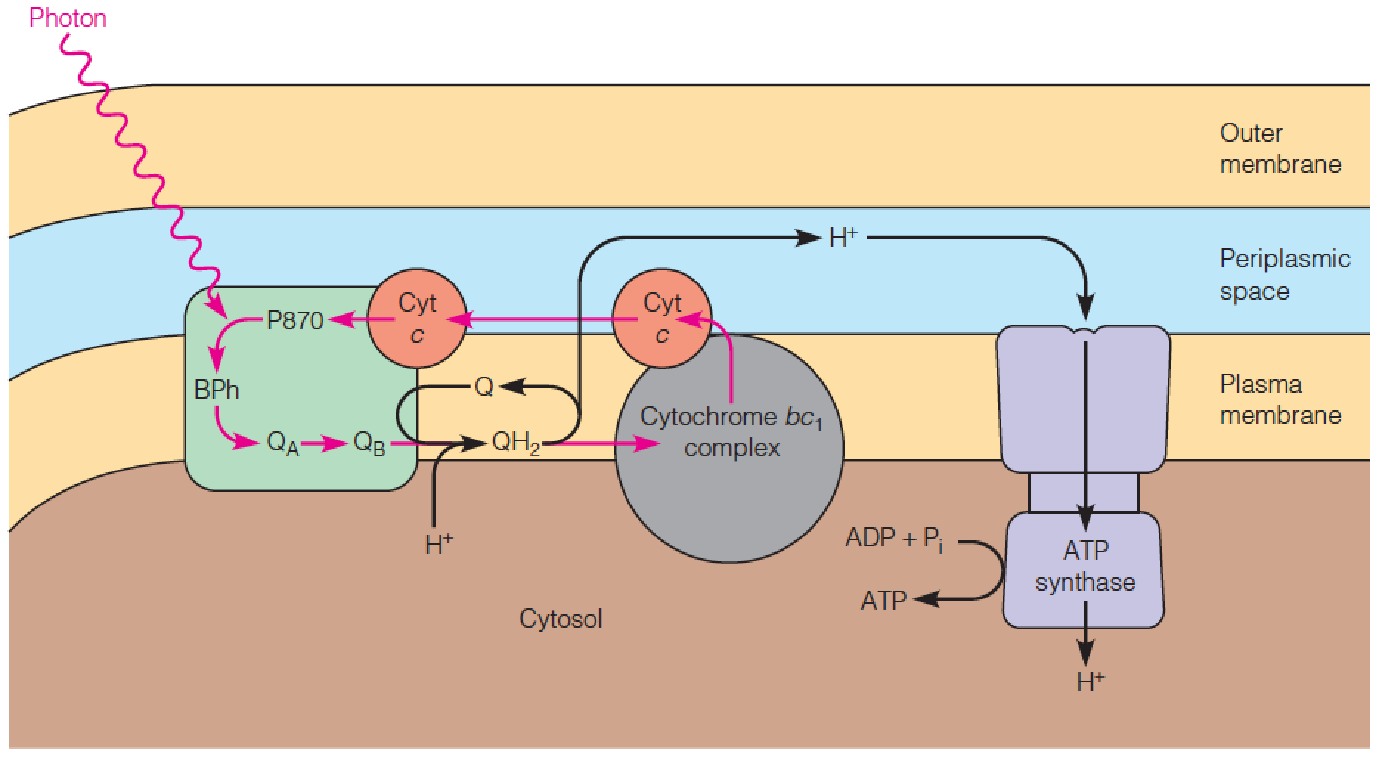
Postulated mechanism for purple bacterial photosynthesis
This process somewhat resembles the photosystem II reactions in the thylakoid, with a reaction center and a membrane-bound cytochrome complex.
However, there is only one kind of reaction center, water is not split, and electron flow is cyclic.
Artificial photosynthesis
In this solar cell, absorbed light energy drives the splitting of water into oxygen and protons, forming H2 fuel.
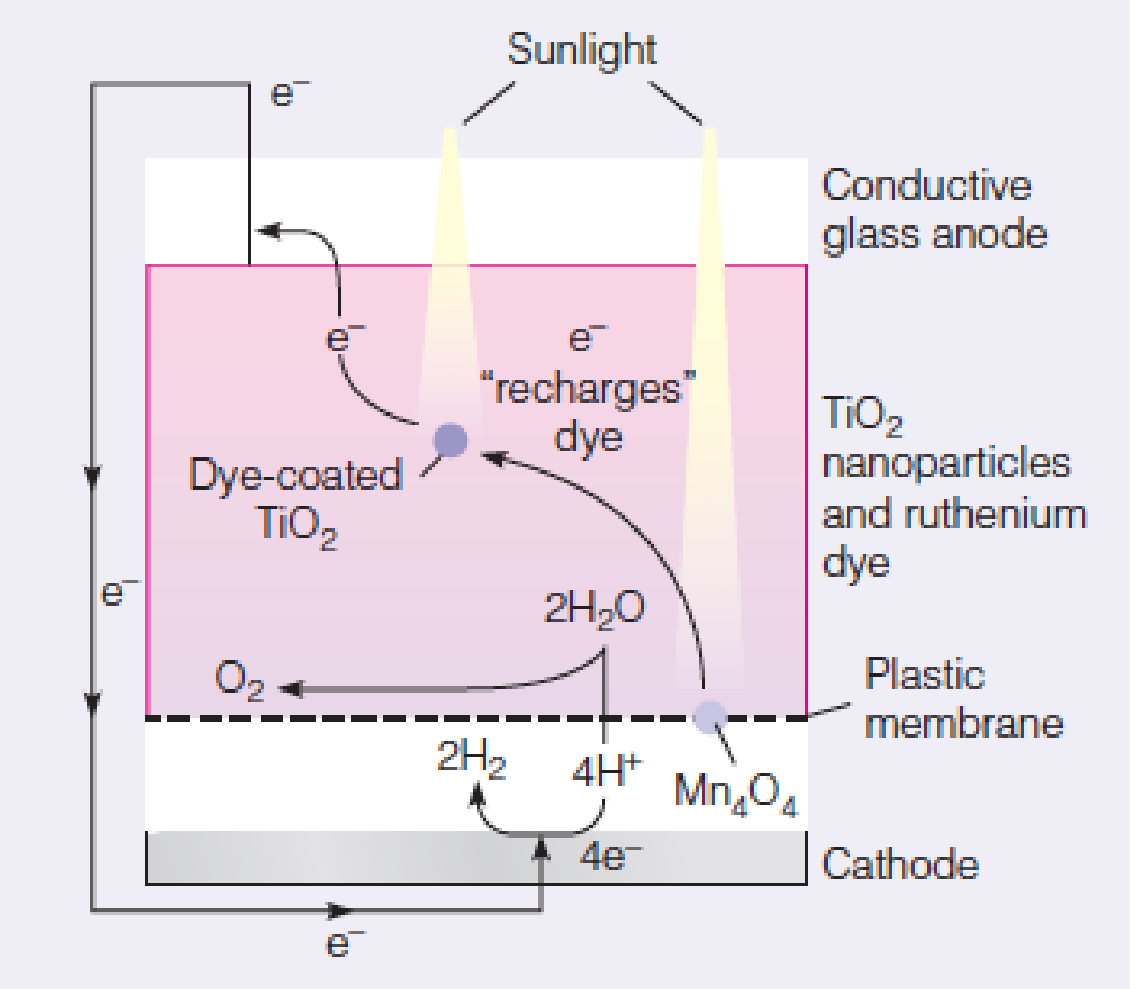
Converts sunlight into chemical fuel (H2)
ATP, NADPH, O2, CO2, Now What?
- Calvin (-Benson) Cycle
- Rubisco (RuBP Carboxylase)
- Photorespiration
四、The Dark Reactions: The Calvin Cycle
Schematic view of the Calvin cycle
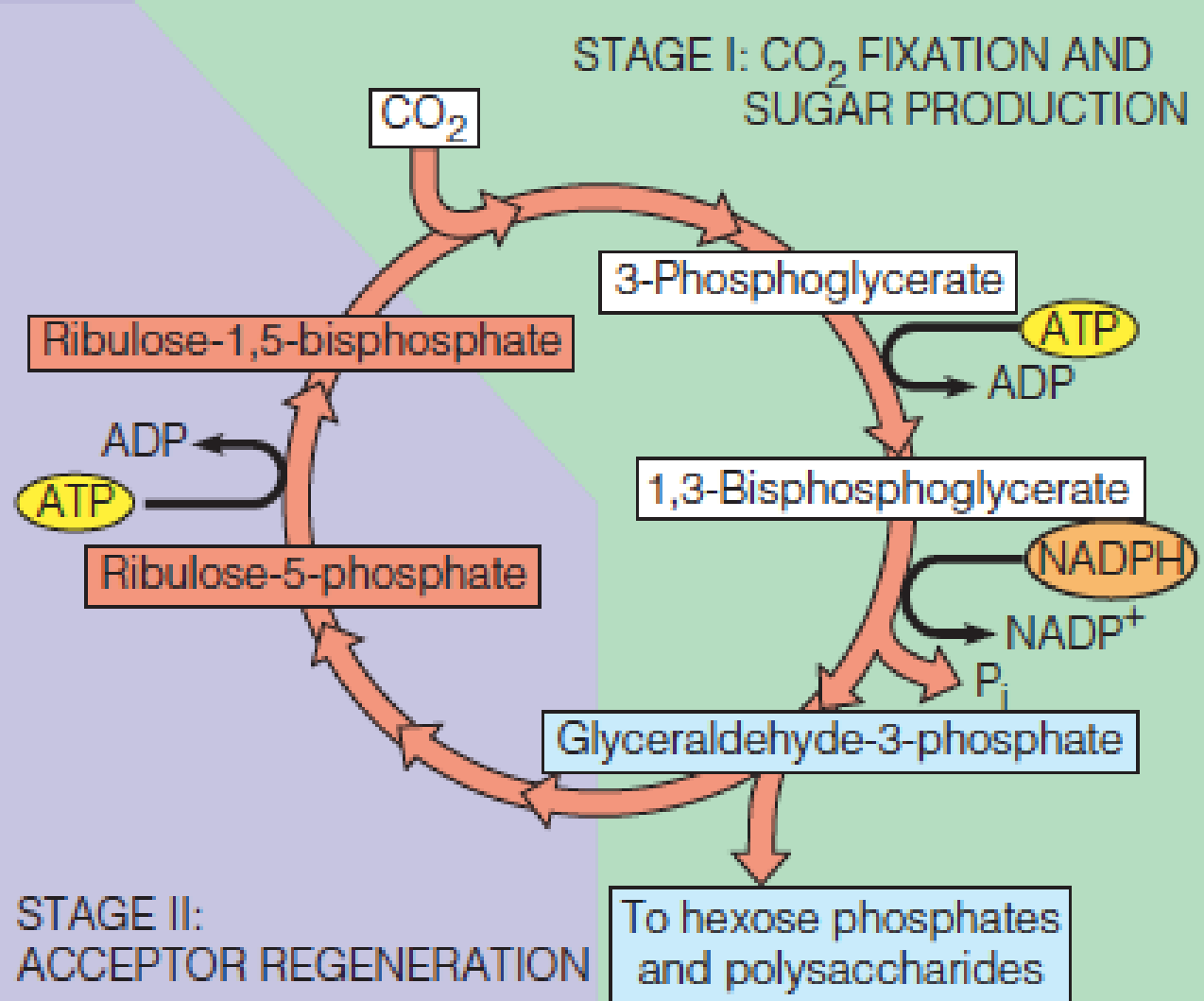
Stage I - CO2 is fixed and glyceraldehyde-3-phosphate (GAP) is produced.
- Part of this GAP is used to make hexose phosphates and eventually polysaccharides.
Stage II - another fraction of the GAP is used to generate the acceptor molecule, ribulose-1,5-BP.
1. Stage I: Carbon Dioxide Fixation and Sugar Production
CO2 is incorporated into glyceraldehyde-3-phosphate (GAP).
The acceptor molecule for CO2 is ribulose-1,5-bisphosphate (RuBP). (1,5-二磷酸核酮糖)
CO2 from the air diffuses into the stroma of the chloroplast, where it is added at the carbonyl carbon of RuBP.
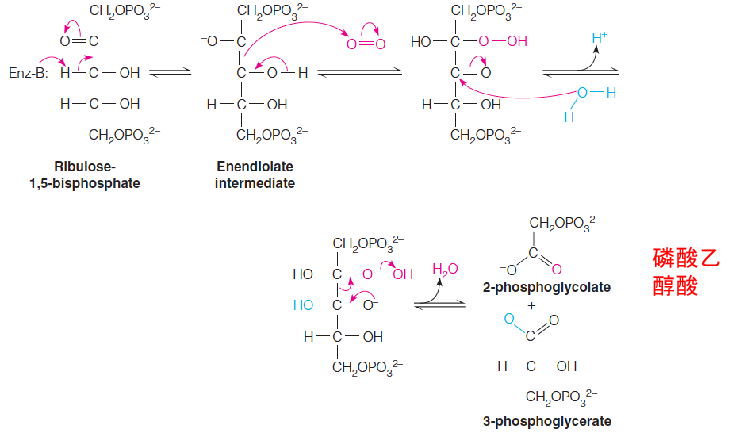
The reaction is catalyzed by the enzyme ribulose-1,5-bisphosphate carboxylase, also known as ribulose-1,5-bisphosphate carboxylase/oxygenase (or rubisco).
This enzyme is one of the most important in the biosphere and certainly the most abundant. It makes up about 15% of all chloroplast proteins, and there are an estimated 40 million tons of it in the world—about 20 pounds for every living person.
The Dark Reactions: The Calvin Cycle
The carboxylase reaction is a complex series of steps in which the actual acceptor is proposed to be the five-carbon enediolate intermediate:
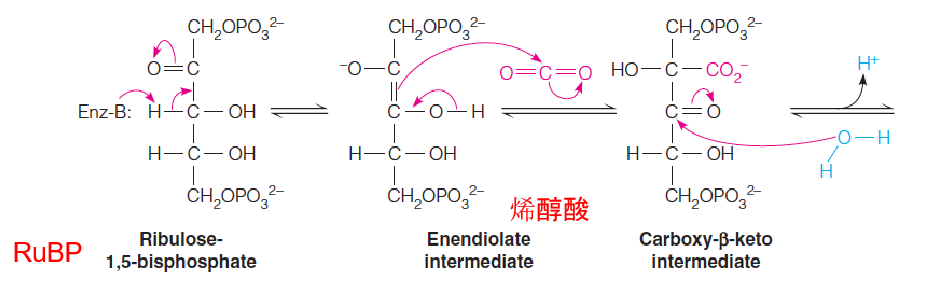
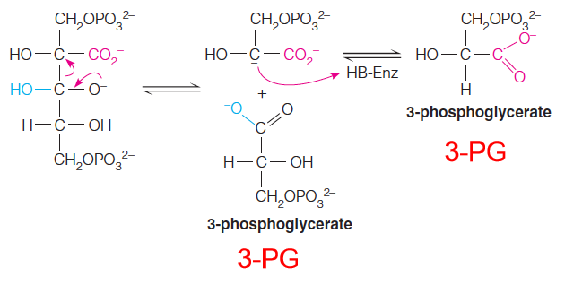
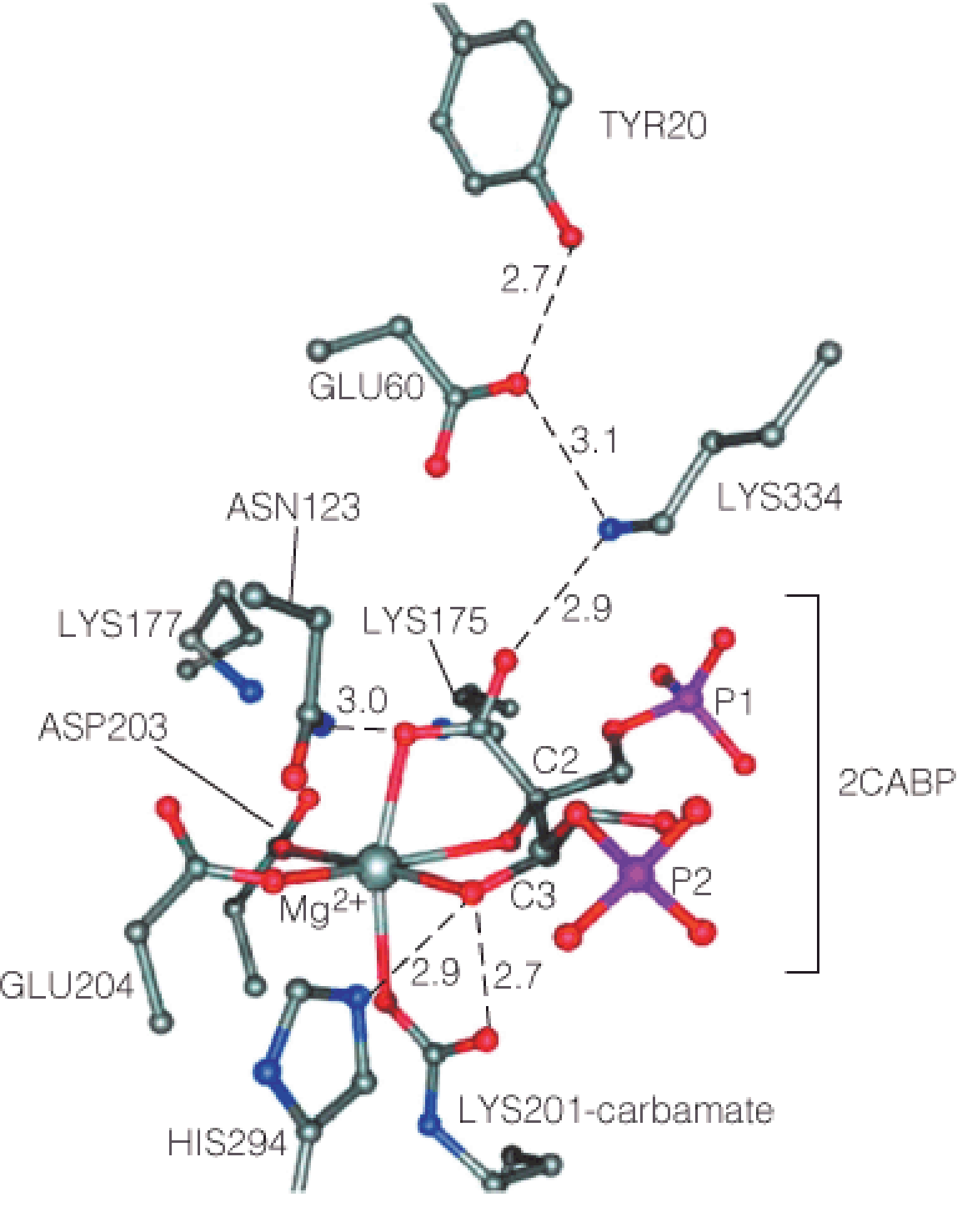
1. Active site of rubisco
This model is based on the X-ray structure of spinach rubisco complexed with Mg2+ and the transition-state analog 2-carboxy-D-arabinitol-1,5-bisphosphate (2CABP).
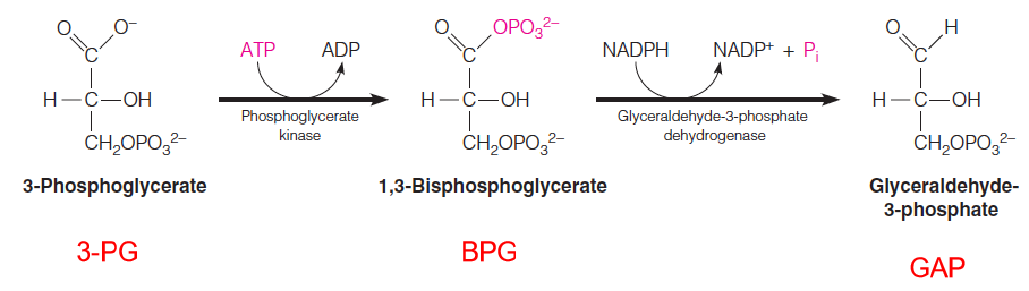
Each molecule of 3PG is phosphorylated by ATP, in a reaction catalyzed by phosphoglycerate kinase.
The 1,3-bisphosphoglycerate so produced is then reduced to glyceraldehyde-3-phosphate (GAP), with accompanying loss of one phosphate.
The reducing agent is NADPH, produced in the light reaction, and the reaction is catalyzed by the enzyme glyceraldehyde-3-phosphate dehydrogenase:
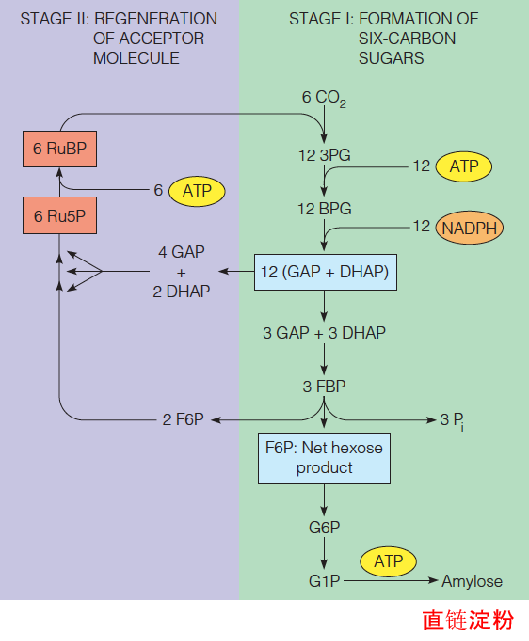
2. Stoichiometry of the Calvin cycle
In six turns of the Calvin cycle, 6 CO2 molecules will have entered and bound to 6 RuBP to yield 12 GAP.
Because GAP is in isomeric equilibrium with DHAP, the 12 GAP may be considered an interconvertible stock of 12 (GAP-DHAP).
Of these, 6 are used to make 3 FBP, of which one constitutes the net hexose product of the 6 turns.
The other 2 FBPs are used, together with the 6 remaining molecules of (GAP-DHAP), to form 6 Ru5P, which are then phosphorylated to regenerate the required 6 RuBP.
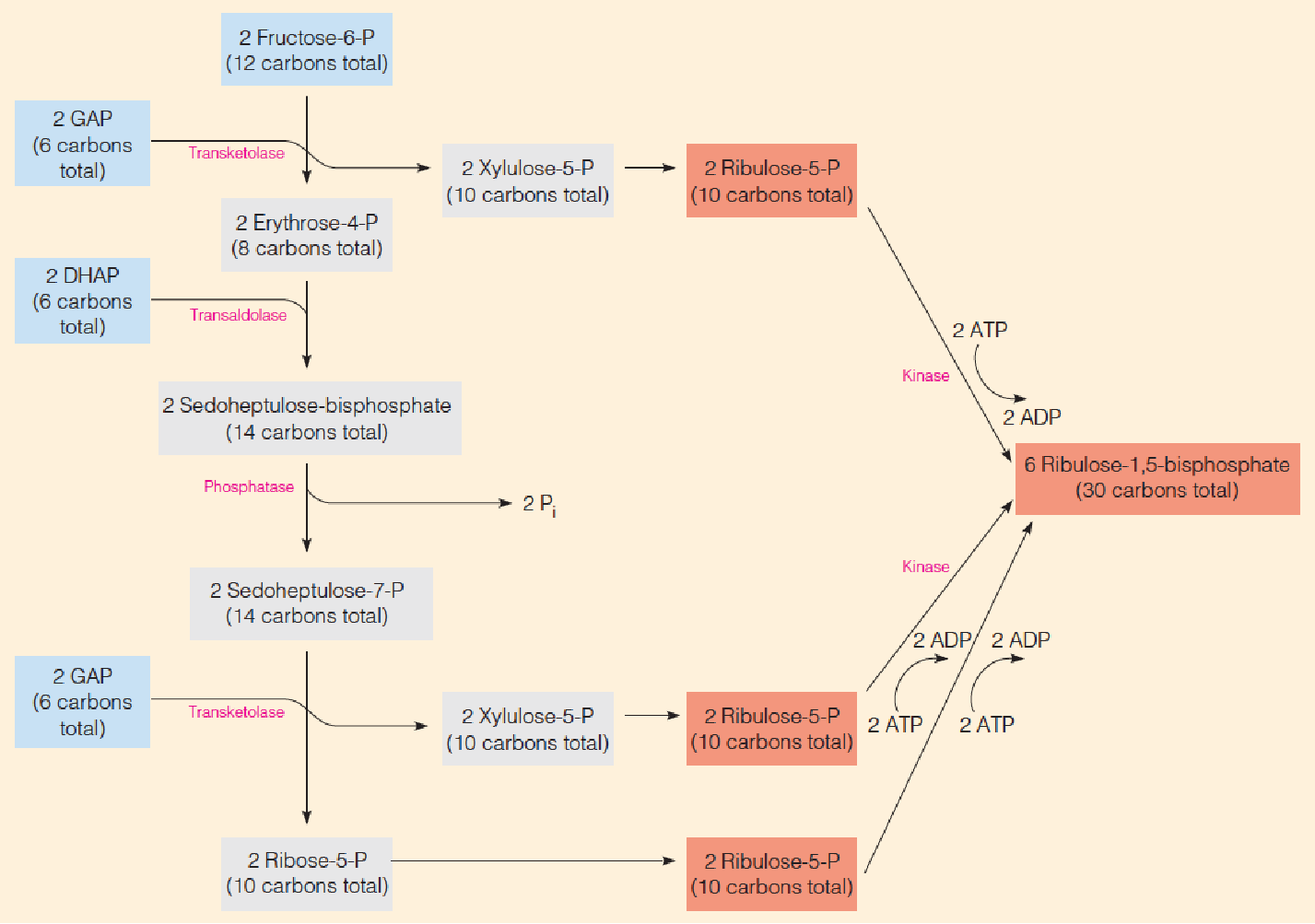
3. Regeneration phase of the Calvin cycle:
Note the similarity of this pathway to parts of the pentose phosphate pathway running in reverse
4. Stage II: Regeneration of the Acceptor
The reactions we have considered to this point can account for the introduction of one carbon into one molecule of hexose, with subsequent formation of oligosaccharides or polysaccharides.
To complete the Calvin cycle, it is necessary to regenerate enough RuBP to keep the cycle going.
This means we will need to regenerate 6 moles of RuBP for every 6 moles of CO2 taken up.
This is accomplished by the set of reactions, which constitute the regenerative phase of the cycle.
Note that the input molecules in this somewhat complex reaction pathway are as follows:
- Two molecules of DHAP and four molecules of GAP, from the six GAP that were diverted to the regeneration pathway.
- Two of the three molecules of fructose-6-phosphate (F6P) that were produced from the remaining three GAP and three DHAP.
五、A Summary of the Light and Dark Reactions in Two-System Photosynthesis
Into & Out of the Dark Reactions
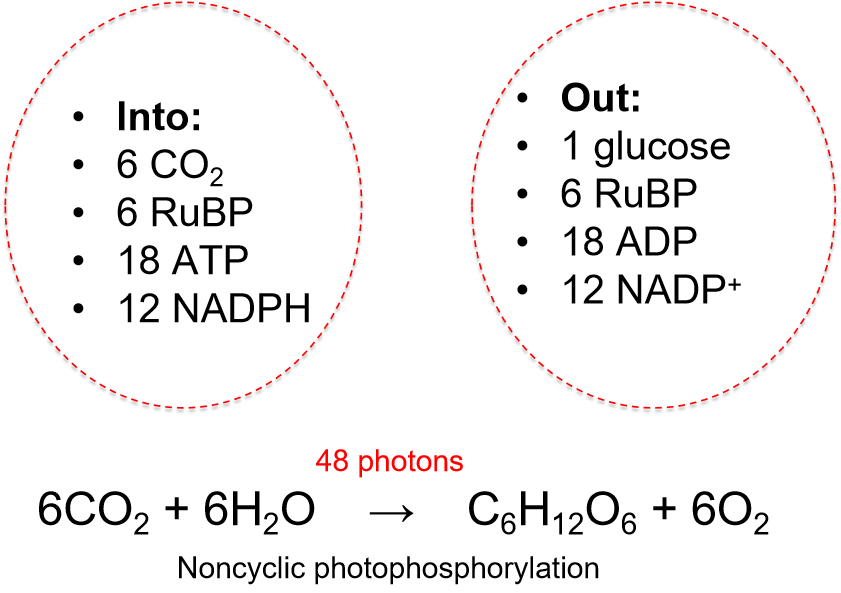
Ribulose bisphosphate carboxylase (rubisco) is a peculiar enzyme.
Under normal environmental conditions, it can behave as an oxygenase as well as a carboxylase.
In the oxygenase reaction, it is proposed that the enediolate intermediate nucleophilically attacks O2 instead of CO2.
- CO2 is low and O2 is high (normal air)
- Km for O2 (21%) is about 10-fold higher than for CO2 (about 0.04%)
- In chloroplast
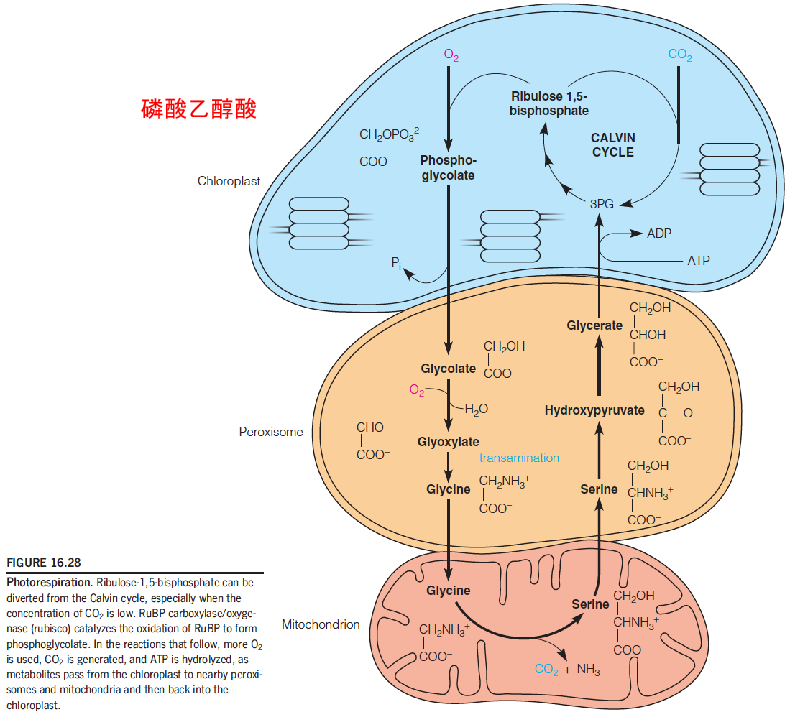
- A wasteful process that costs ATP and limits the carbon assimilation by the plant (“flaw design” :
- RuBP is lost from the Calvin cycle;
- The fixation of CO2 is reversed;
- O2 is consumed;CO2 is released
- Only a part (75%) of the carbon is returned to the chloroplast
- ATP is expended
- Evolution can only work with what it has at hand (about 3 billion yrs ago: O2 is very low, oxygenation activity is negligible).
- As O2 levels (21%) increased, oxygenase activity became significant).
- Modify the rubisco during evolution? Not much
- Can we engineer an more efficient Rubisco
- Enzyme? Not successful yet.
六、Photorespiration and the C4 Cycle
The C4 Cycle
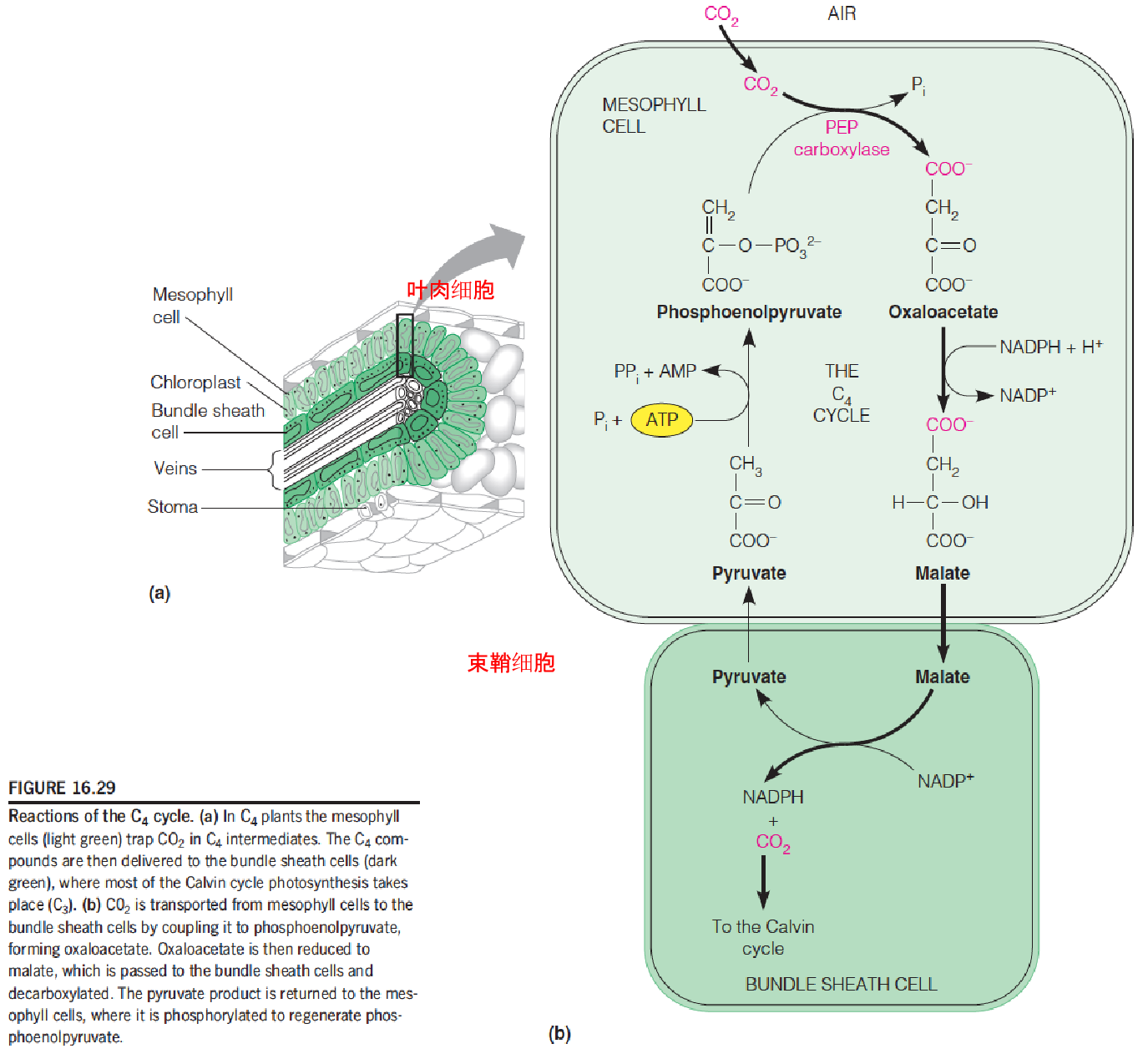
C4 plants: a plant that utilizes the C4 carbon fixation pathway in which the CO2 is first bound to a phosphoenolpyruvate in mesophyll cell (叶肉细胞) resulting in the formation of four-carbon compound (oxaloacetate) that is shuttled to the bundle sheath cell where it will be decarboxylated to liberate the CO2 to be utilized in the C3 pathway.
C4 plants (maize, sugarcane):
- tropical plants;
- High illumination, high temperature, CO2 deletion;
- CO2 concentrating mechanism(浓缩了二氧化碳)
- Help fix CO2 from photorespiration
- PEP carboxylase: no oxygenase activity(避免了光呼吸的低效率); low Km for CO2
七、Evolution of Photosynthesis
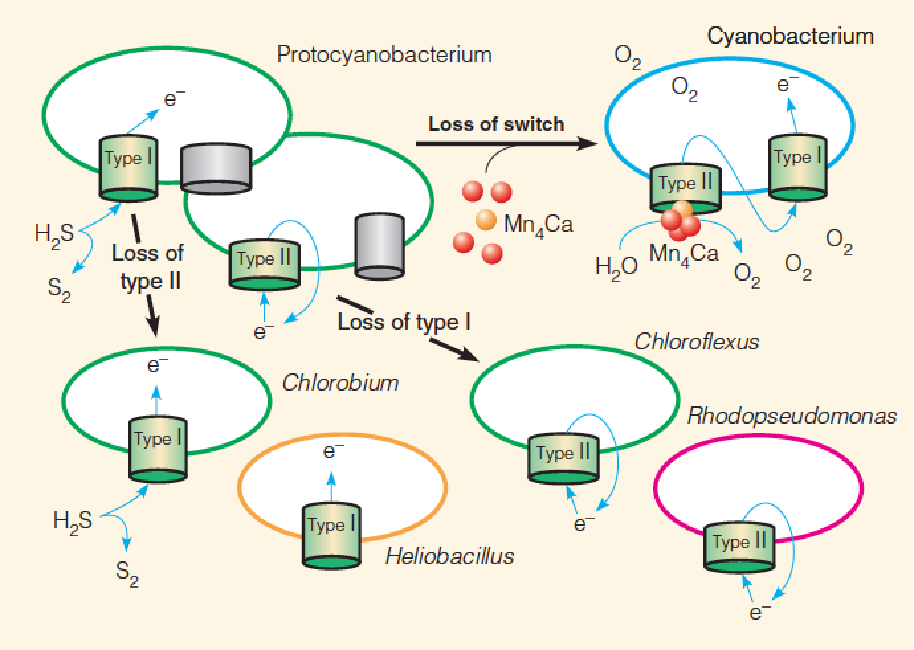
Proposed evolution of oxygenic photosynthesis: